I gangli autonomi sono formati da neuroni. Gangli vegetativi. Un numero così grande di neuroni non può essere innervato da una singola fibra pregangliare.
Il tessuto nervoso (con la partecipazione di una serie di altri tessuti) forma il sistema nervoso, che garantisce la regolazione di tutti i processi vitali nel corpo e la sua interazione con l'ambiente esterno.
Anatomicamente, il sistema nervoso è diviso in centrale e periferico. Quello centrale comprende il cervello e il midollo spinale, quello periferico unisce i nodi nervosi, i nervi e le terminazioni nervose.
Il sistema nervoso si sviluppa dal tubo neurale e dalla placca gangliare. Il cervello e gli organi di senso si differenziano dalla parte cranica del tubo neurale. Dalla parte del tronco del tubo neurale - il midollo spinale, dalla placca gangliare si formano i nodi spinali e autonomi e il tessuto cromaffino del corpo.
Nervi (gangli)
I nodi nervosi, o gangli, sono gruppi di neuroni al di fuori del sistema nervoso centrale. Assegna nodi nervosi sensibili e autonomi.
I gangli sensoriali si trovano lungo le radici posteriori del midollo spinale e lungo il decorso dei nervi cranici. I neuroni afferenti nella spirale e nei gangli vestibolari sono bipolari, nei restanti gangli sensoriali sono pseudo-unipolari.
ganglio spinale (ganglio spinale)
Il ganglio spinale ha una forma fusiforme, circondato da una capsula di tessuto connettivo denso. Dalla capsula, sottili strati di tessuto connettivo penetrano nel parenchima del nodo, in cui si trovano i vasi sanguigni.
I neuroni del ganglio spinale sono caratterizzati da un grande corpo sferico e un nucleo leggero con un nucleolo ben visibile. Le cellule sono disposte in gruppi, principalmente lungo la periferia dell'organo. Il centro del ganglio spinale è costituito principalmente da processi di neuroni e strati sottili di endoneurio che trasportano i vasi sanguigni. I dendriti delle cellule nervose vanno come parte della parte sensibile dei nervi spinali misti alla periferia e terminano lì con i recettori. Gli assoni formano collettivamente le radici posteriori che portano gli impulsi nervosi al midollo spinale o al midollo allungato.
Nei nodi spinali dei vertebrati superiori e dell'uomo, i neuroni bipolari diventano pseudounipolari durante la maturazione. Un singolo processo parte dal corpo di un neurone pseudounipolare, che avvolge ripetutamente la cellula e spesso forma un groviglio. Questo processo si divide a forma di T in rami afferenti (dendritici) ed efferenti (assonali).
I dendriti e gli assoni delle cellule nel nodo e oltre sono ricoperti da guaine mieliniche di neurolemmociti. Il corpo di ciascuna cellula nervosa nel ganglio spinale è circondato da uno strato di cellule di oligodendroglia appiattite, qui chiamate gliociti del mantello, o gliociti gangliari, o cellule satelliti. Si trovano intorno al corpo del neurone e hanno piccoli nuclei arrotondati. All'esterno, la guaina gliale del neurone è ricoperta da una sottile guaina fibrosa di tessuto connettivo. Le cellule di questo guscio si distinguono per la forma ovale dei nuclei.
I neuroni gangliari spinali contengono neurotrasmettitori come acetilcolina, acido glutammico, sostanza P.
Nodi autonomi (vegetativi).
I nodi nervosi autonomi si trovano:
lungo la colonna vertebrale (gangli paravertebrali);
davanti alla colonna vertebrale (gangli prevertebrali);
nella parete degli organi: cuore, bronchi, tubo digerente, vescica (gangli intramurali);
vicino alla superficie di questi organi.
Le fibre pregangliari mieliniche contenenti processi di neuroni del sistema nervoso centrale si avvicinano ai nodi vegetativi.
In base alle loro caratteristiche funzionali e localizzazione, i nodi nervosi autonomi sono divisi in simpatici e parasimpatici.
La maggior parte degli organi interni ha una doppia innervazione autonomica, cioè riceve fibre postgangliari da cellule situate nei nodi simpatici e parasimpatici. Le risposte mediate dai loro neuroni hanno spesso la direzione opposta (ad esempio, la stimolazione simpatica migliora l'attività cardiaca, mentre la stimolazione parasimpatica la inibisce).
Il piano generale della struttura dei nodi vegetativi è simile. All'esterno, il nodo è ricoperto da una sottile capsula di tessuto connettivo. I nodi vegetativi contengono neuroni multipolari, caratterizzati da una forma irregolare, un nucleo posizionato eccentricamente. Spesso ci sono neuroni multinucleati e poliploidi.
Ogni neurone e i suoi processi sono circondati da una guaina di cellule satelliti gliali: i gliociti del mantello. La superficie esterna della membrana gliale è ricoperta da una membrana basale, al di fuori della quale è presente una sottile membrana di tessuto connettivo.
A causa della loro elevata autonomia, della complessità dell'organizzazione e delle caratteristiche del metabolismo mediatore, i gangli intramurali degli organi interni e le vie ad essi associate si distinguono talvolta in una divisione metasimpatica indipendente del sistema nervoso autonomo.
Nei nodi intramurali, l'istologo russo Dogel A.S. sono descritti tre tipi di neuroni:
cellule efferenti di tipo I ad assone lungo;
cellule afferenti di uguale lunghezza di tipo II;
cellule associative di tipo III.
I neuroni efferenti dell'assone lungo (cellule di Dogel di tipo I) sono neuroni numerosi e grandi con dendriti corti e un lungo assone che esce dal nodo all'organo di lavoro, dove forma terminazioni motorie o secretorie.
I neuroni afferenti a crescita uguale (cellule Dogel di tipo II) hanno lunghi dendriti e un assone che si estende oltre questo nodo in quelli vicini. Queste cellule fanno parte degli archi riflessi locali come legame recettoriale, che vengono chiusi senza che un impulso nervoso entri nel sistema nervoso centrale.
I neuroni associativi (cellule Dogel di tipo III) sono neuroni intercalari locali che collegano diverse cellule di tipo I e II con i loro processi.
I neuroni dei gangli nervosi autonomi, come quelli dei nodi spinali, sono di origine ectodermica e si sviluppano dalle cellule della cresta neurale.
nervi periferici
I nervi, o tronchi nervosi, collegano i centri nervosi del cervello e del midollo spinale con i recettori e gli organi di lavoro o con i nodi nervosi. I nervi sono formati da fasci di fibre nervose, che sono unite da guaine di tessuto connettivo.
La maggior parte dei nervi sono misti, cioè comprendono le fibre nervose afferenti ed efferenti.
I fasci nervosi contengono fibre mieliniche e non mieliniche. Il diametro delle fibre e il rapporto tra fibre nervose mielinizzate e non mielinizzate in nervi diversi non sono gli stessi.
Sulla sezione trasversale del nervo sono visibili sezioni dei cilindri assiali delle fibre nervose e delle membrane gliali che le rivestono. Alcuni nervi contengono singole cellule nervose e piccoli gangli.
Tra le fibre nervose nella composizione del fascio nervoso ci sono sottili strati di tessuto connettivo fibroso lasso - endoneurium. Ci sono poche cellule al suo interno, predominano le fibre reticolari, passano piccoli vasi sanguigni.
Fasci separati di fibre nervose sono circondati da perineurio. Il perineurio è costituito da strati alternati di cellule densamente impaccate e sottili fibre di collagene orientate lungo il nervo.
Il guscio esterno del tronco nervoso - l'epinevrio - è un tessuto connettivo fibroso denso ricco di fibroblasti, macrofagi e cellule adipose. Contiene vasi sanguigni e linfatici, terminazioni nervose sensibili
istologia privata.
Istologia privata- la scienza della struttura microscopica e dell'origine degli organi. Ogni organo è composto da 4 tessuti.
Organi del sistema nervoso.
Su base funzionale
1. sistema nervoso somatico- partecipa all'innervazione del corpo umano e all'attività nervosa superiore.
un. Dipartimento centrale:
io. Midollo spinale - nuclei delle corna posteriori e anteriori
ii. Cervello - corteccia cerebellare ed emisferi cerebrali
b. Dipartimento periferico:
io. gangli spinali
ii. gangli cranici
iii. tronchi nervosi
2. sistema nervoso autonomo- svolge il lavoro degli organi interni, innerva i miociti lisci e rappresenta i nervi secretori.
1) comprensivo:
un. Dipartimento centrale:
io. Midollo spinale - nuclei delle corna laterali della regione toraco-lombare
ii. cervello - ipotalamo
b. Dipartimento periferico:
io. Gangli simpatici
ii. tronchi nervosi
2) Parasimpatico:
un. Dipartimento centrale:
io. Midollo spinale - nuclei delle corna laterali della regione sacrale
ii. Cervello - nuclei del tronco cerebrale, ipotalamo
b. Dipartimento periferico:
io. gangli parasimpatici
ii. tronchi nervosi
iii. Gangli spinali e cranici
Anatomicamente Gli organi del sistema nervoso si dividono in:
1. Sistema nervoso periferico.
2. Sistema nervoso centrale.
Fonti embrionali di sviluppo:
1. neuroectoderma(dà origine al parenchima degli organi).
2. mesenchima(dà origine allo stroma degli organi, un insieme di strutture ausiliarie che assicurano il funzionamento del parenchima).
Gli organi del sistema nervoso funzionano in relativo isolamento dall'ambiente, separandosi da esso. barriere biologiche. Tipi di barriere biologiche:
1. Ematoneurale (delimita il sangue dai neuroni).
2. Liquoroneurale (delimita il liquido cerebrospinale dai neuroni).
3. Ematoliquore (delimita il liquido cerebrospinale dal sangue).
Funzioni del sistema nervoso:
1. Regolazione delle funzioni dei singoli organi interni.
2. Integrazione degli organi interni nei sistemi di organi.
3. Garantire la relazione dell'organismo con l'ambiente esterno.
4. Garantire una maggiore attività nervosa.
Tutte le funzioni sono basate sul principio riflesso. La base materiale è arco riflesso, composto da 3 link: afferente, associativo e efferente. Sono distribuiti ai singoli organi del sistema nervoso.
Organi del sistema nervoso periferico:
1. Tronchi nervosi (nervi).
2. Nodi nervosi (gangli).
3. Terminazioni nervose.
tronchi nervosi - si tratta di fasci di fibre nervose, unite da un sistema di membrane di tessuto connettivo. I tronchi nervosi sono misti, cioè ciascuno ha fibre mieliniche e amieliniche, a seguito delle quali vengono serviti i sistemi nervosi somatico e autonomo.
La struttura del tronco nervoso:
1. parenchima: fibre nervose amieliniche e mielinizzate + microgangli.
2. Stroma: membrane del tessuto connettivo:
1) Perineurio(guaine perineurali: RVNST + vasi sanguigni + ependimogliociti + liquido cerebrospinale).
2) epinevrio(PVNST + vasi sanguigni).
3) Perineurio(scissione dall'epinevrio nel tronco).
4) Endoneurio(RVNST + vasi sanguigni).
C'è uno spazio simile a una fessura nel perineurio - guaina perineurale a fessura che è riempito liquore(fluido biologico circolante). Componenti strutturali delle pareti della guaina perineurale:
1. Ependimogliociti prismatici bassi.
2. Membrana basale.
3. Placca subependimale.
4. Vasi sanguigni.
Il liquore nella vagina perineurale può essere assente. A volte vengono iniettati anestetici, antibiotici (perché la malattia si diffonde attraverso di loro).
Funzioni dei tronchi nervosi:
1. Conduzione (condurre un impulso nervoso).
2. Trofico (nutrizionale).
4. Sono il collegamento iniziale nella secrezione e nella circolazione del liquido cerebrospinale.
Rigenerazione dei tronchi nervosi:
1. Rigenerazione fisiologica(ripristino molto attivo delle membrane a causa dei fibroblasti).
2. Rigenerazione riparativa(viene ripristinata quella sezione del tronco nervoso, le cui fibre nervose non hanno perso il contatto con il pericarion - sono in grado di crescere di 1 mm / giorno; i segmenti periferici delle fibre nervose non vengono ripristinati).
Nervi (gangli) - gruppi o cooperazioni di neuroni, prelevati dal cervello. I nodi nervosi sono "vestiti" in capsule.
Tipi di gangli:
1. Spinale.
2. cranico.
3. Vegetativo.
gangli spinali - ispessimenti sulle sezioni iniziali delle radici posteriori del midollo spinale; questo è un gruppo di neuroni afferenti (sensibili) (sono i primi neuroni nella catena dell'arco riflesso).
La struttura del ganglio spinale:
1. Stroma:
1) capsula di tessuto connettivo esterno, composta da 2 fogli:
un. foglio esterno (tessuto connettivo denso - continuazione dell'epineurio del nervo spinale)
b. foglio interno (multi-tessuto: RVNST, gliociti; analogo del perineurio del nervo spinale; ci sono fessure che passano ai setti intraorgano, riempiti di liquido cerebrospinale).
2) partizioni intraorgano che si estendono dalla capsula al nodo
b. vasi sanguigni e linfatici
c. fibre nervose
d. terminazioni nervose
3) proprie capsule di tessuto connettivo di neuroni pseudounipolari
un. tessuto connettivo fibroso
b. epitelio ependimogliale squamoso a strato singolo
c. spazio perineuronale con liquido cerebrospinale
2. parenchima:
1) la parte centrale (fibre nervose mielinizzate - processi di neuroni pseudo-unipolari)
2) parte periferica (neuroni pseudounipolari + gliociti del mantello (oligodendrogliociti)).
Funzioni del ganglio spinale:
1. Partecipazione all'attività riflessa (i primi neuroni nel circuito dell'arco riflesso).
2. Sono il collegamento iniziale nel trattamento delle informazioni afferenti.
3. Funzione barriera (barriera ematoneurale).
4. Sono un collegamento nella circolazione del liquido cerebrospinale.
Fonti di sviluppo embrionale del ganglio spinale:
1. Piastra gangliare (dà origine agli elementi del parenchima dell'organo).
2. Mesenchima (dà origine agli elementi dello stroma dell'organo).
Gangli del sistema nervoso autonomo - situato dopo il midollo spinale, partecipa alla creazione di archi autonomici.
Tipi di gangli del sistema nervoso autonomo:
1. comprensivo:
1) paravertebrale;
2) Prevertebrale;
2. Parasimpatico:
1) Intraorganico (intramurale);
2) Perioorganico (paraorganico);
3) Nodi vegetativi della testa (lungo i nervi cranici).
La struttura dei gangli del sistema nervoso autonomo:
1. Stroma: la struttura è simile allo stroma del ganglio spinale.
2.1. Parenchima dei gangli simpatici: neuroni localizzati casualmente in tutto il ganglio + cellule satelliti + capsula del tessuto connettivo.
1) neuroni adrenergici efferenti multipolari ad assone lungo
2) piccoli neuroni associativi multipolari equidistanti adrenergici intensamente fluorescenti (MIF)
3) fibre colinergiche mieliniche pregangliari (assoni dei neuroni delle corna laterali del midollo spinale)
4) fibre nervose adrenergiche non mielinizzate postgangliari (assoni dei grandi neuroni gangliari)
5) fibre nervose associative intragangliari non mielinizzate (assoni del MITO - neuroni).
2.2. Parenchima dei gangli parasimpatici:
1) neuroni colinergici efferenti multipolari ad assone lungo (Dogel tipo I).
2) neuroni colinergici afferenti multipolari dendritici lunghi (tipo Dogel II): dendrite - al recettore, assone - ai tipi 1 e 3.
3) neuroni colinergici associativi multipolari equidistanti (Dogel tipo III).
4) fibre nervose colinergiche mieliniche pregangliari (assoni delle corna laterali del midollo spinale).
5) fibre nervose colinergiche non mielinizzate postgangliari (assoni dei neuroni di tipo I Dogel).
Funzioni dei gangli del sistema nervoso autonomo:
1. comprensivo:
1) Condurre gli impulsi agli organi di lavoro (2.1.1)
2) Propagazione dell'impulso all'interno del ganglio (effetto frenante) (2.1.2)
2. Parasimpatico:
1) Condurre un impulso agli organi di lavoro (2.2.1)
2) Condurre un impulso dagli interocettori all'interno di archi riflessi locali (2.2.2)
3) propagazione dell'impulso all'interno o tra i gangli (2.2.3).
Fonti di sviluppo embrionale dei gangli del sistema nervoso autonomo:
1. Placca gangliare (neuroni e neuroglia).
2. Mesenchima (tessuto connettivo, vasi sanguigni).
È un sistema di tessuti e organi costruiti da tessuto nervoso. Evidenzia:
Regione centrale: cervello e midollo spinale
Periferico: gangli autonomi e sensoriali, nervi periferici, terminazioni nervose.
C'è anche una divisione in:
Reparto somatico (animale, cerebrospinale);
Reparto vegetativo (autonomo): parti simpatiche e parasimpatiche.
Il sistema nervoso è formato dalle seguenti fonti embrionali: tubo neurale, cresta neurale (placca gangliare) e placode embrionali. Gli elementi tissutali delle membrane sono derivati mesenchimali. Nella fase di chiusura del neuroporo, l'estremità anteriore del tubo si espande in modo significativo, le pareti laterali si ispessiscono, formando l'inizio di tre vescicole cerebrali. La vescica che giace cranialmente forma il proencefalo, la vescica media forma il mesencefalo e il cervello posteriore (romboidale) si sviluppa dalla terza vescica, che passa nell'anlage del midollo spinale. Subito dopo, il tubo neurale si piega quasi ad angolo retto e, attraverso i solchi di restringimento, la prima vescica è divisa nelle sezioni finali e intermedie e la terza vescica cerebrale nel midollo allungato e nelle sezioni posteriori del cervello. I derivati delle vescicole cerebrali medie e posteriori formano il tronco cerebrale e sono formazioni antiche; mantengono il principio segmentale della struttura, che scompare nei derivati del diencefalo e del telencefalo. In quest'ultimo si concentrano le funzioni integrative. È così che si formano cinque parti del cervello: finale e diencefalo, medio, midollo allungato e romboencefalo (nell'uomo, ciò si verifica approssimativamente alla fine della 4a settimana di sviluppo embrionale). Il telencefalo forma i due emisferi del cervello.
Nell'istogenesi embrionale e nell'organogenesi del sistema nervoso, lo sviluppo di diverse parti del cervello avviene a velocità diverse (eterocrone). In precedenza si formano le parti caudali del sistema nervoso centrale (midollo spinale, tronco encefalico); il tempo della formazione finale delle strutture cerebrali varia notevolmente. In alcune parti del cervello ciò si verifica dopo la nascita (cervelletto, ippocampo, bulbo olfattivo); in ogni parte del cervello ci sono gradienti spazio-temporali nella formazione di popolazioni neuronali che formano una struttura unica del centro nervoso.
Il midollo spinale è una parte del sistema nervoso centrale, nella cui struttura sono più chiaramente conservate le caratteristiche degli stadi embrionali di sviluppo del cervello dei vertebrati: la natura tubolare della struttura e la segmentazione. Nelle sezioni laterali del tubo neurale, la massa delle cellule aumenta rapidamente, mentre le sue parti dorsale e ventrale non aumentano di volume e mantengono il loro carattere ependimale. Le pareti laterali ispessite del tubo neurale sono divise da un solco longitudinale nella dorsale - alare e ventrale - la piastra principale. In questa fase di sviluppo, si possono distinguere tre zone nelle pareti laterali del tubo neurale: ependima che riveste il canale centrale, intermedia (strato di mantello) e marginale (velo marginale). Dallo strato del mantello si sviluppa successivamente la materia grigia del midollo spinale e dal velo marginale la sua sostanza bianca. I neuroblasti delle colonne anteriori si differenziano in motoneuroni (motoneuroni) dei nuclei delle corna anteriori. I loro assoni escono dal midollo spinale e formano le radici anteriori dei nervi spinali. Nelle colonne posteriori e nella zona intermedia si sviluppano vari nuclei di cellule intercalari (associative). I loro assoni, che entrano nella sostanza bianca del midollo spinale, fanno parte di vari fasci conduttori. Le corna posteriori comprendono i processi centrali dei neuroni sensoriali dei nodi spinali.
Contemporaneamente allo sviluppo del midollo spinale, vengono posati i nodi spinali e periferici del sistema nervoso autonomo. Il materiale di partenza per loro sono gli elementi cellulari staminali della cresta neurale, che, attraverso differenziazioni divergenti, si sviluppano nelle direzioni neuroblastica e glioblastica. Parte delle cellule della cresta neurale migrano alla periferia verso i siti di localizzazione dei nodi del sistema nervoso autonomo, dei paragangli, delle cellule neuroendocrine della serie APUD e del tessuto cromaffino.
Sistema nervoso periferico.
Il sistema nervoso periferico combina nodi nervosi periferici, tronchi e terminazioni.
I gangli nervosi (nodi) - strutture formate da ammassi di neuroni al di fuori del sistema nervoso centrale - sono divisi in sensibili e autonomi (vegetativi). I gangli sensoriali contengono neuroni afferenti pseudounipolari o bipolari (nei gangli spirali e vestibolari) e si trovano principalmente lungo le radici posteriori del midollo spinale (nodi sensoriali dei nervi spinali) e alcuni nervi cranici. I gangli sensoriali dei nervi spinali sono fusiformi e ricoperti da una capsula di denso tessuto connettivo fibroso. Alla periferia del ganglio ci sono densi ammassi di corpi di neuroni pseudo-unipolari e la parte centrale è occupata dai loro processi e da sottili strati di endoneurio situati tra di loro, che trasportano vasi. I gangli del nervo autonomo sono formati da gruppi di neuroni multipolari, sui quali numerose sinapsi formano fibre pregangliari - processi di neuroni i cui corpi si trovano nel SNC.
Nervo. Costruzione e rigenerazione. Gangli spinali. Caratteristiche morfofunzionali.
I nervi (tronchi nervosi) collegano i centri nervosi del cervello e del midollo spinale con i recettori e gli organi di lavoro. Sono formati da fasci di fibre mieliniche e non mieliniche, che sono unite da componenti del tessuto connettivo (gusci): endoneurio, perinevrio ed epinevrio. La maggior parte dei nervi sono misti, cioè comprendono fibre afferenti ed efferenti.
Endoneurium - strati sottili di tessuto connettivo fibroso lasso con piccoli vasi sanguigni, che circondano le singole fibre nervose e le collegano in un unico fascio. Il perineurio è una guaina che copre ciascun fascio di fibre nervose dall'esterno ed estende le partizioni in profondità nel fascio. Ha una struttura lamellare e immagini di strati concentrici di cellule appiattite simili a fibroblasti collegate da articolazioni dense e scanalate. Tra gli strati di cellule negli spazi pieni di liquido, ci sono componenti della membrana basale e fibre di collagene orientate longitudinalmente. L'epinevrio è la guaina esterna del nervo che lega insieme i fasci di fibre nervose. È costituito da un denso tessuto connettivo fibroso contenente cellule adipose, vasi sanguigni e linfatici.
Midollo spinale. Caratteristiche morfofunzionali. Sviluppo. Struttura della materia grigia e bianca. composizione neurale.
Il midollo spinale è costituito da due metà simmetriche, delimitate anteriormente da una profonda fessura mediana e posteriormente da un setto di tessuto connettivo. La parte interna dell'organo è più scura: questa è la sua materia grigia. Alla periferia del midollo spinale c'è una sostanza bianca più chiara. La materia grigia del midollo spinale è costituita dai corpi dei neuroni, dalle fibre mieliniche amieliniche e sottili e dalla neuroglia. Il componente principale della materia grigia, che la distingue dal bianco, sono i neuroni multipolari. Le sporgenze della materia grigia sono chiamate corna. Ci sono corna anteriori o ventrali, posteriori o dorsali e laterali o laterali. Durante lo sviluppo del midollo spinale, i neuroni si formano dal tubo neurale, raggruppati in 10 strati o in placche. Caratteristica di una persona
la seguente architettura delle tavole indicate: le tavole I-V corrispondono alle corna posteriori, le tavole VI-VII - alla zona intermedia, le tavole VIII-IX - alle corna anteriori, la tavola X - alla zona del canale quasi centrale. La materia grigia del cervello è costituita da tre tipi di neuroni multipolari. Il primo tipo di neuroni è filogeneticamente più antico ed è caratterizzato da pochi dendriti lunghi, diritti e debolmente ramificati (tipo isodendritico). Il secondo tipo di neuroni ha un gran numero di dendriti fortemente ramificati che si intrecciano formando "grovigli" (tipo idiodendritico). Il terzo tipo di neuroni, in termini di grado di sviluppo dei dendriti, occupa una posizione intermedia tra il primo e il secondo tipo. La sostanza bianca del midollo spinale è un insieme di fibre prevalentemente mielinizzate orientate longitudinalmente. I fasci di fibre nervose che comunicano tra le diverse parti del sistema nervoso sono chiamati vie del midollo spinale.
Cervello. Fonti di sviluppo. Caratteristiche morfofunzionali generali degli emisferi cerebrali. Organizzazione neuronale degli emisferi cerebrali. Cito- e mieloarchitettura della corteccia cerebrale. Cambiamenti legati all'età nella corteccia.
Nel cervello si distinguono la materia grigia e quella bianca, ma la distribuzione di queste due componenti è molto più complicata qui che nel midollo spinale. La maggior parte della materia grigia del cervello si trova sulla superficie del cervello e nel cervelletto, formando la loro corteccia. Una parte più piccola forma numerosi nuclei del tronco cerebrale.
Struttura. La corteccia cerebrale è rappresentata da uno strato di materia grigia. È più fortemente sviluppato nel giro centrale anteriore. L'abbondanza di solchi e convoluzioni aumenta notevolmente l'area della materia grigia del cervello. Le sue varie parti, che differiscono tra loro per alcune caratteristiche della posizione e della struttura delle cellule (citoarchitettonica), la posizione delle fibre (mieloarchitettura) e significato funzionale, sono chiamati campi. Sono luoghi di maggiore analisi e sintesi degli impulsi nervosi. nettamente definito
non ci sono confini tra di loro. La corteccia è caratterizzata dalla disposizione di cellule e fibre a strati. Lo sviluppo della corteccia cerebrale umana (neocorteccia) nell'embriogenesi avviene dalla zona germinale ventricolare del telencefalo, dove si trovano cellule proliferanti scarsamente specializzate. I neurociti neocorticali si differenziano da queste cellule. In questo caso, le cellule perdono la loro capacità di dividersi e migrare verso la placca corticale emergente. Innanzitutto, i neurociti dei futuri strati I e VI entrano nella placca corticale, ad es. gli strati più superficiali e profondi della corteccia. Quindi i neuroni degli strati V, IV, III e II vengono incorporati nella direzione dall'interno e verso l'esterno. Questo processo viene eseguito a causa della formazione di cellule in piccole aree della zona ventricolare in diversi periodi di embriogenesi (eterocrona). In ciascuna di queste aree si formano gruppi di neuroni, allineati in sequenza lungo una o più fibre.
glia radiale a forma di colonna.
Citoarchitettura della corteccia cerebrale. I neuroni multipolari della corteccia hanno una forma molto diversa. Tra questi ci sono i neuroni piramidali, stellati, fusiformi, aracnidi e orizzontali. I neuroni della corteccia si trovano in strati non nettamente delimitati. Ogni strato è caratterizzato dalla predominanza di un qualsiasi tipo di cellula. Nella zona motoria della corteccia si distinguono 6 strati principali: I - molecolare, II - granulare esterno, III - neuroni nuramidali, IV - granulare interno, V - gangliare, VI - strato di cellule polimorfiche. Lo strato molecolare della corteccia contiene un piccolo numero di piccole cellule associative a forma di fuso. I loro neuriti corrono paralleli alla superficie del cervello come parte del plesso tangenziale delle fibre nervose dello strato molecolare. Lo strato granulare esterno è formato da piccoli neuroni che hanno una forma arrotondata, angolare e piramidale e neurociti stellati. I dendriti di queste cellule salgono nello strato molecolare. I neuriti entrano nella sostanza bianca o, formando archi, entrano anche nel plesso tangenziale delle fibre dello strato molecolare. Lo strato più largo della corteccia cerebrale è quello piramidale. Dalla parte superiore della cellula piramidale parte il dendrite principale, che si trova nello strato molecolare. Il neurite della cellula piramidale si allontana sempre dalla sua base. Lo strato granulare interno è formato da piccoli neuroni stellati. È costituito da un gran numero di fibre orizzontali. Lo strato gangliare della corteccia è formato da grandi piramidi e la regione del giro precentrale contiene piramidi giganti.
Lo strato di cellule polimorfiche è formato da neuroni di varie forme.
Mieloarchitettura della corteccia. Tra le fibre nervose della corteccia cerebrale, si possono individuare le fibre associative che collegano singole parti della corteccia di un emisfero, le fibre commissurali che collegano la corteccia di diversi emisferi e le fibre di proiezione, sia afferenti che efferenti, che collegano la corteccia con i nuclei delle parti inferiori della centrale
sistema nervoso.
Cambiamenti di età. Al 1 ° anno di vita si osservano la tipizzazione della forma dei neuroni piramidali e stellati, il loro aumento, lo sviluppo di arborizzazione dendritica e assonale e le connessioni intraensemble lungo la verticale. All'età di 3 anni, negli insiemi vengono rivelati gruppi di neuroni "nidificati", fasci dendritici verticali più chiaramente formati e fasci di fibre radianti. All'età di 5-6 anni, il polimorfismo neuronale aumenta; il sistema di connessioni intra-insieme lungo l'orizzontale diventa più complicato a causa della crescita in lunghezza e ramificazione dei dendriti laterali e basali dei neuroni piramidali e dello sviluppo dei terminali laterali dei loro dendriti apicali. All'età di 9-10 anni, i gruppi cellulari aumentano, la struttura dei neuroni ad assoni corti diventa molto più complicata e la rete di collaterali assonici di tutte le forme di interneuroni si espande. All'età di 12-14 anni, le forme specializzate di neuroni piramidali sono chiaramente contrassegnate negli insiemi; tutti i tipi di interneuroni raggiungono un alto livello di differenziazione. All'età di 18 anni, l'organizzazione d'insieme della corteccia, in termini di parametri principali della sua architettura, raggiunge il livello di quella degli adulti.
Cervelletto. Struttura e caratteristiche morfofunzionali. Composizione neuronale della corteccia cerebellare, gliociti. Connessioni interneuronali.
Cervelletto. È l'organo centrale dell'equilibrio e della coordinazione dei movimenti. È collegato al tronco cerebrale da fasci conduttori afferenti ed efferenti, che insieme formano tre paia di peduncoli cerebellari. Ci sono molte convoluzioni e solchi sulla superficie del cervelletto, che ne aumentano significativamente l'area. Sul taglio vengono creati solchi e convoluzioni
caratteristico per l'immagine del cervelletto dell '"albero della vita". La maggior parte della materia grigia nel cervelletto si trova in superficie e ne forma la corteccia. Una parte più piccola della materia grigia si trova in profondità nella sostanza bianca sotto forma di nuclei centrali. Al centro di ogni giro c'è uno strato sottile
materia bianca, ricoperta da uno strato di materia grigia: la corteccia. Nella corteccia cerebellare si distinguono tre strati: quello esterno è lo strato molecolare, quello intermedio è lo strato gangliare, o lo strato di neuroni a forma di pera, e quello interno è granulare. Lo strato gangliare contiene neuroni a forma di pera. Hanno neuriti, che, lasciando la corteccia cerebellare, formano il collegamento iniziale del suo efferente
percorsi di frenata. Dal corpo a forma di pera, 2-3 dendriti si estendono nello strato molecolare, che penetrano per l'intero spessore dello strato molecolare. Dalla base dei corpi di queste cellule partono i neuriti, che passano attraverso lo strato granulare della corteccia cerebellare nella sostanza bianca e terminano sulle cellule dei nuclei cerebellari. Lo strato molecolare contiene due tipi principali di neuroni: canestro e stellato. I neuroni canestro si trovano nel terzo inferiore dello strato molecolare. I loro lunghi e sottili dendriti si ramificano principalmente su un piano situato trasversalmente al giro. I lunghi neuriti delle cellule corrono sempre attraverso il giro e parallelamente alla superficie sopra i neuroni a forma di pera. I neuroni stellati si trovano sopra le cellule canestro e sono di due tipi. I piccoli neuroni stellati sono dotati di sottili dendriti corti e neuriti debolmente ramificati che formano sinapsi. I grandi neuroni stellati hanno dendriti e neuriti lunghi e altamente ramificati. strato granuloso. Il primo tipo di cellule in questo strato può essere considerato neuroni granulari o cellule granulari. La cellula ha 3-4 dendriti corti,
terminando nello stesso strato con rami terminali a forma di zampa d'uccello. I neuriti delle cellule granulari passano nello strato molecolare e in esso sono divisi in due rami, orientati parallelamente alla superficie della corteccia lungo il giro del cervelletto. Il secondo tipo di cellule nello strato granulare del cervelletto sono grandi neuroni stellati inibitori. Esistono due tipi di tali cellule: con neuriti corti e lunghi. I neuroni con neuriti corti si trovano vicino allo strato gangliare. I loro dendriti ramificati si diffondono nello strato molecolare e formano sinapsi con fibre parallele - assoni di cellule granulari. I neuriti vengono inviati allo strato granulare ai glomeruli del cervelletto e terminano in sinapsi ai rami terminali dei dendriti delle cellule granulari.
Alcuni neuroni stellati con neuriti lunghi hanno dendriti e neuriti abbondantemente ramificati nello strato granulare, che si estendono nella sostanza bianca. Il terzo tipo di cellule sono cellule orizzontali a forma di fuso. Hanno un piccolo corpo allungato, da cui si estendono lunghi dendriti orizzontali in entrambe le direzioni, terminando negli strati gangliari e granulari. I neuriti di queste cellule danno collaterali allo strato granulare e vanno a
sostanza bianca. Gliociti. La corteccia cerebellare contiene vari elementi gliali. Lo strato granulare contiene astrociti fibrosi e protoplasmatici. I peduncoli dei processi fibrosi degli astrociti formano membrane perivascolari. Tutti gli strati del cervelletto contengono oligodendrociti. Lo strato granulare e la sostanza bianca del cervelletto sono particolarmente ricchi di queste cellule. Le cellule gliali con nuclei scuri si trovano nello strato gangliare tra i neuroni a forma di pera. I processi di queste cellule vengono inviati alla superficie della corteccia e formano fibre gliali dello strato molecolare del cervelletto. Connessioni interneuronali. Le fibre afferenti che entrano nella corteccia cerebellare sono rappresentate da due tipi: le fibre muscose e le cosiddette fibre rampicanti. Le fibre muscose vanno come parte dei tratti oliva-cerebellare e cerebellopontino e indirettamente attraverso le cellule dei granuli hanno un effetto eccitante sulle cellule a forma di pera.
Le fibre rampicanti entrano nella corteccia cerebellare, apparentemente, lungo le vie dorso-cerebellare e vestibolocerebellare. Attraversano lo strato granulare, si uniscono ai neuroni a forma di pera e si diffondono lungo i loro dendriti, terminando sulla loro superficie con sinapsi. Le fibre rampicanti trasmettono l'eccitazione direttamente ai neuroni piriformi.
Sistema nervoso autonomo (vegetativo). Caratteristiche morfofunzionali generali. Dipartimenti. Struttura dei gangli extramurali ed intramurali.
L'ANS è diviso in simpatico e parasimpatico. Entrambi i sistemi prendono simultaneamente parte all'innervazione degli organi e hanno l'effetto opposto su di essi. È costituito dalle sezioni centrali, rappresentate dai nuclei della sostanza grigia del cervello e del midollo spinale, e da quelle periferiche: tronchi nervosi, nodi (gangli) e plessi.
A causa della loro elevata autonomia, della complessità dell'organizzazione e delle caratteristiche del metabolismo del mediatore, i gangli intramurali e le vie ad essi associate si distinguono in un dipartimento metasimpatico indipendente del NS autonomo. Esistono tre tipi di neuroni:
Neuroni efferenti ad assone lungo (cellule di Dogel tipo I) con dendriti corti e un lungo assone che si estende oltre il nodo fino alle cellule dell'organo di lavoro, su cui forma terminazioni motorie o secretorie.
I neuroni afferenti a crescita uguale (cellule di Dogel di tipo II) contengono lunghi dendriti e un assone che si estende oltre questo ganglio in quelli vicini e forma sinapsi sulle cellule di tipo I e III. Fanno parte degli archi riflessi locali come collegamento recettoriale, che vengono chiusi senza che un impulso nervoso entri nel sistema nervoso centrale.
Le cellule associative (cellule Dogel di tipo III) sono neuroni intercalari locali che collegano diverse cellule di tipo I e II con i loro processi. I dendriti di queste cellule non vanno oltre il nodo e gli assoni vanno ad altri nodi, formando sinapsi su cellule di tipo I.
Sistema nervoso effettua l'unificazione di parti del corpo in un unico insieme (integrazione), assicura la regolazione di vari processi, il coordinamento delle funzioni di vari organi e tessuti e l'interazione del corpo con l'ambiente esterno. Percepisce diverse informazioni provenienti dall'ambiente esterno e dagli organi interni, le elabora e genera segnali che forniscono risposte adeguate agli stimoli attivi. L'attività del sistema nervoso si basa su archi riflessi- catene di neuroni che forniscono reazioni organi di lavoro (organi bersaglio) in risposta alla stimolazione del recettore. Negli archi riflessi, i neuroni collegati tra loro da sinapsi formano tre collegamenti: recettore (afferente), effettore e tra loro associativo (inserire).
Dipartimenti del sistema nervoso
Divisione anatomica dei dipartimenti sistema nervoso:
(1)sistema nervoso centrale (SNC) -
include testa e dorsale cervello;
(2)sistema nervoso periferico - include gangli nervosi periferici (nodi), nervi e terminazioni nervose(descritto nella sezione "Tessuto nervoso").
Divisione fisiologica dei dipartimenti del sistema nervoso(a seconda della natura dell'innervazione di organi e tessuti):
(1)sistema nervoso somatico (animale). - controlla principalmente le funzioni del movimento volontario;
(2)sistema nervoso autonomo (vegetativo). - regola l'attività degli organi interni, vasi e ghiandole.
Il sistema nervoso autonomo è diviso nell'interazione tra loro comprensivo e divisioni parasimpatiche, che differiscono per la localizzazione dei nodi e dei centri periferici nel cervello, nonché per la natura dell'effetto sugli organi interni.
Il sistema nervoso somatico e autonomo comprende collegamenti situati nel sistema nervoso centrale e nel sistema nervoso periferico. Tessuto funzionalmente all'avanguardia gli organi del sistema nervoso sono tessuto nervoso, compresi neuroni e glia. I cluster di neuroni nel SNC sono comunemente chiamati core, e nel sistema nervoso periferico gangli (nodi). Sono chiamati fasci di fibre nervose nel sistema nervoso centrale tratti, nella periferica nervi.
Nervi(tronchi nervosi) collegano i centri nervosi del cervello e del midollo spinale con i recettori e gli organi di lavoro. Sono formati in fasci mielina e fibre nervose amieliniche che sono uniti da componenti del tessuto connettivo (gusci): endoneurio, perinevrio e epinevrio(Fig. 114-118). La maggior parte dei nervi sono misti, cioè includono fibre nervose afferenti ed efferenti.
Endoneurio - strati sottili di tessuto connettivo fibroso lasso con piccoli vasi sanguigni, che circondano le singole fibre nervose e le collegano in un unico fascio.
Perineurio - una guaina che copre ciascun fascio di fibre nervose dall'esterno e fornisce partizioni in profondità nel fascio. Ha una struttura lamellare ed è formato da strati concentrici di cellule appiattite simili a fibroblasti collegate da giunzioni strette e gap. Tra gli strati di cellule negli spazi pieni di liquido, ci sono componenti della membrana basale e fibre di collagene orientate longitudinalmente.
epinevrio - la guaina esterna del nervo che lega insieme fasci di fibre nervose. È costituito da un denso tessuto connettivo fibroso contenente cellule adipose, vasi sanguigni e linfatici (vedi Fig. 114).
Strutture nervose rivelate da vari metodi di colorazione. Vari metodi di colorazione istologica consentono uno studio più dettagliato e selettivo dei singoli componenti
nervo. Così, osmizzazione dà una colorazione a contrasto delle guaine mieliniche delle fibre nervose (consentendo di valutarne lo spessore e differenziare tra fibre mieliniche e non mieliniche), ma i processi dei neuroni e dei componenti del tessuto connettivo del nervo rimangono molto debolmente colorati o non colorati (vedi Fig. 114 e 115). Quando dipingi ematossilina-eosina le guaine mieliniche non sono colorate, i processi dei neuroni hanno una colorazione leggermente basofila, tuttavia i nuclei dei neurolemmociti nelle fibre nervose e tutti i componenti del tessuto connettivo del nervo sono ben rilevati (vedi Fig. 116 e 117). In colorato con nitrato d'argento i processi dei neuroni sono colorati in modo brillante; le guaine mieliniche rimangono non colorate, i componenti del tessuto connettivo del nervo sono scarsamente rilevati, la loro struttura non viene tracciata (vedi Fig. 118).
Gangli nervosi (nodi)- le strutture formate da cluster di neuroni al di fuori del SNC - sono divise in sensibile e autonomo(vegetativo). I gangli sensoriali contengono neuroni afferenti pseudounipolari o bipolari (nei gangli spirali e vestibolari) e si trovano principalmente lungo le radici posteriori del midollo spinale (nodi sensoriali dei nervi spinali) e alcuni nervi cranici.
Gangli sensoriali (nodi) dei nervi spinali a forma di fuso e coperto capsula di tessuto connettivo fibroso denso. Alla periferia del ganglio ci sono densi ammassi di corpi neuroni pseudounipolari, e la parte centrale è occupata dai loro processi e da sottili strati di endoneurium situati tra loro, che portano vasi (Fig. 121).
Neuroni sensoriali pseudounipolari sono caratterizzati da un corpo sferico e da un nucleo leggero con nucleolo ben visibile (Fig. 122). Il citoplasma dei neuroni contiene numerosi mitocondri, cisterne del reticolo endoplasmatico granulare, elementi del complesso del Golgi (vedi Fig. 101), lisosomi. Ogni neurone è circondato da uno strato di cellule oligodendroglia appiattite adiacenti ad esso. o gliociti del mantello) con piccoli nuclei arrotondati; all'esterno della membrana gliale è presente una sottile capsula di tessuto connettivo (vedi Fig. 122). Un processo parte dal corpo di un neurone pseudounipolare, dividendosi a forma di T in rami periferici (afferenti, dendritici) e centrali (efferenti, assonali), che sono ricoperti da guaine mieliniche. processo periferico(ramo afferente) termina con recettori,
processo centrale(ramo efferente) come parte della radice posteriore entra nel midollo spinale (vedi Fig. 119).
Gangli del nervo autonomo formato da gruppi di neuroni multipolari, sui quali si formano numerose sinapsi fibre pregangliari- processi di neuroni i cui corpi si trovano nel sistema nervoso centrale (vedi Fig. 120).
Classificazione dei gangli autonomi. Per localizzazione: i gangli possono essere localizzati lungo la colonna vertebrale (gangli paravertebrali) o davanti a lui (gangli prevertebrali) così come nella parete degli organi: cuore, bronchi, tubo digerente, vescica, ecc. (gangli intramurali- vedi, ad esempio, fig. 203, 209, 213, 215) o in prossimità della loro superficie.
Funzionalmente, i gangli nervosi autonomi sono divisi in simpatici e parasimpatici. Questi gangli differiscono per la loro localizzazione (simpatica giacciono para- e prevertebrale, parasimpatico - intramurale o vicino agli organi), nonché per la localizzazione dei neuroni che danno origine alle fibre pregangliari, la natura dei neurotrasmettitori e la direzione delle reazioni mediate dalle loro cellule. La maggior parte degli organi interni ha una doppia innervazione autonomica. Il piano generale della struttura dei gangli dei nervi simpatici e parasimpatici è simile.
La struttura dei gangli autonomi. Il ganglio autonomo è ricoperto esternamente da tessuto connettivo. capsula e contiene corpi diffusi o raggruppati neuroni multipolari, i loro processi sotto forma di fibre ed endoneurium non mielinizzate o (raramente) mielinizzate (Fig. 123). I corpi dei neuroni sono basofili, di forma irregolare, contengono un nucleo posizionato eccentricamente; ci sono cellule multinucleari e poliploidi. I neuroni sono circondati (di solito in modo incompleto) da guaine di cellule gliali (cellule gliali satellitari, o gliociti del mantello). Al di fuori della membrana gliale c'è una sottile membrana di tessuto connettivo (Fig. 124).
gangli intramurali e i percorsi ad essi associati, per la loro elevata autonomia, la complessità dell'organizzazione e le peculiarità dello scambio dei mediatori, si distinguono da alcuni autori come un divisione metasimpatica sistema nervoso autonomo. Tre tipi di neuroni sono descritti nei gangli intramurali (vedi Fig. 120):
1) Neuroni efferenti ad assone lungo (cellule Dogel di tipo I) con dendriti corti e un lungo assone che si estende oltre il nodo
alle cellule dell'organo di lavoro, su cui forma terminazioni motorie o secretorie.
2)Neuroni afferenti a crescita uguale (cellule Dogel di tipo II) contengono lunghi dendriti e un assone che si estende oltre il ganglio dato in quelli vicini e forma sinapsi su cellule di tipo I e III. Fanno parte degli archi riflessi locali come collegamento recettoriale, che vengono chiusi senza che un impulso nervoso entri nel sistema nervoso centrale.
3)Cellule di associazione (cellule di Dogel di tipo III)- neuroni intercalari locali, che connettono diverse cellule di tipo I e II con i loro processi. I dendriti di queste cellule non vanno oltre il nodo e gli assoni vanno ad altri nodi, formando sinapsi su cellule di tipo I.
Archi riflessi nelle parti somatica (animale) e autonomica (vegetativa) del sistema nervoso hanno una serie di caratteristiche (vedi Fig. 119 e 120). Le principali differenze sono nei collegamenti associativi ed effettori, poiché il collegamento del recettore è simile: è formato da neuroni pseudounipolari afferenti, i cui corpi si trovano nei gangli sensoriali. I processi periferici di queste cellule formano le terminazioni nervose sensoriali, mentre i processi centrali entrano nel midollo spinale come parte delle radici posteriori.
Collegamento associativo nell'arco somatico è rappresentato dai neuroni intercalari, i cui dendriti e corpi si trovano in corna posteriori del midollo spinale e gli assoni vanno a corna anteriori, trasmettere impulsi ai corpi e ai dendriti dei neuroni efferenti. Nell'arco autonomo si trovano i dendriti e i corpi dei neuroni intercalari corna laterali del midollo spinale e gli assoni (fibre pregangliari) lasciano il midollo spinale come parte delle radici anteriori, dirigendosi verso uno dei gangli autonomi, dove terminano sui dendriti e sui corpi dei neuroni efferenti.
Collegamento effettore nell'arco somatico è formato da motoneuroni multipolari, i cui corpi e dendriti si trovano nelle corna anteriori del midollo spinale, e gli assoni lasciano il midollo spinale come parte delle radici anteriori, vanno al ganglio sensoriale e quindi, come parte del nervo misto, al muscolo scheletrico, sulle cui fibre i loro rami formano sinapsi neuromuscolari. Nell'arco autonomo, il collegamento effettore è formato da neuroni multipolari, i cui corpi si trovano nei gangli autonomi e gli assoni (fibre postgangliari) come parte dei tronchi nervosi e dei loro rami vengono inviati alle cellule degli organi di lavoro - muscoli lisci, ghiandole, cuore.
Organi del sistema nervoso centrale Midollo spinale
Midollo spinale ha l'aspetto di un cordone tondeggiante, espanso nelle regioni cervicale e lombosacrale e penetrato dal canale centrale. Si compone di due metà simmetriche, divise davanti fessura mediana anteriore, dietro - solco mediano posteriore ed è caratterizzato da una struttura segmentale; ad ogni segmento è associata una coppia anteriore (motore, ventrale) e una coppia indietro (sensibile, radici dorsali). Nel midollo spinale ci sono Materia grigia, situato nella sua parte centrale, e sostanza bianca, sdraiato alla periferia (Fig. 125).
materia grigia sulla sezione ha l'aspetto di una farfalla (vedi Fig. 125) e comprende accoppiati anteriore (ventrale), posteriore (dorsale) e corna laterali (laterali). Le corna della materia grigia di entrambe le parti simmetriche del midollo spinale sono collegate tra loro nell'area commessure grigie anteriori e posteriori. La materia grigia contiene i corpi, i dendriti e (in parte) gli assoni dei neuroni, nonché le cellule gliali. Tra i corpi dei neuroni c'è neuropillo- una rete formata da fibre nervose e processi di cellule gliali. I neuroni si trovano nella materia grigia sotto forma di ammassi che non sono sempre nettamente delimitati. (noccioli).
Le corna posteriori contengono diversi nuclei formati interneuroni multipolari, su cui terminano gli assoni delle cellule pseudounipolari dei gangli sensibili (vedi Fig. 119), così come le fibre delle vie discendenti dai centri sopraspinali che giacciono sopra. Gli assoni dei neuroni intercalari a) terminano nella sostanza grigia del midollo spinale sui motoneuroni che si trovano nelle corna anteriori (vedi Fig. 119); b) formare connessioni intersegmentali all'interno della sostanza grigia del midollo spinale; c) escono nella sostanza bianca del midollo spinale, dove formano vie ascendenti e discendenti (tratti).
Le corna laterali, ben espresse a livello dei segmenti toracico e sacrale del midollo spinale, contengono nuclei formati dai corpi neuroni intercalari multipolari, che appartengono alle divisioni simpatiche e parasimpatiche del sistema nervoso autonomo (vedi Fig. 120). Sui dendriti e sui corpi di queste cellule terminano gli assoni: a) neuroni pseudo-unipolari che trasportano impulsi da recettori situati negli organi interni, b) neuroni dei centri di regolazione delle funzioni autonome, i cui corpi si trovano nel midollo allungato. Gli assoni dei neuroni autonomi, lasciando il midollo spinale come parte delle radici anteriori, formano un
fibre gliniche che portano ai nodi simpatici e parasimpatici.
Le corna anteriori contengono motoneuroni multipolari (motoneuroni), combinati in nuclei, ciascuno dei quali di solito si estende in più segmenti. Ci sono grandi motoneuroni α e più piccoli motoneuroni γ sparsi tra di loro. Sui processi e sui corpi dei motoneuroni sono presenti numerose sinapsi che hanno effetti eccitatori e inibitori su di esse. Sulla fine dei motoneuroni: collaterali dei processi centrali di gabbie pseudounipolari di nodi sensoriali; neuroni intercalari, i cui corpi giacciono nelle corna posteriori del midollo spinale; assoni di piccoli neuroni intercalari locali (cellule di Renshaw) associati a collaterali di assoni di motoneuroni; fibre delle vie discendenti dei sistemi piramidale ed extrapiramidale, che trasportano gli impulsi dalla corteccia cerebrale e dai nuclei del tronco cerebrale. I corpi dei motoneuroni contengono grossi grumi di sostanza cromatofila (vedi Fig. 100) e sono circondati da gliociti (Fig. 126). Gli assoni dei motoneuroni lasciano il midollo spinale radici anteriori, inviato al ganglio sensibile e poi, come parte del nervo misto, al muscolo scheletrico, sulle cui fibre si formano sinapsi neuromuscolari(vedi fig. 119).
Canale centrale (vedi Fig. 128) passa al centro della materia grigia ed è circondato davanti e punte grigie posteriori(vedi fig. 125). È riempito di liquido cerebrospinale ed è rivestito da un unico strato di cellule ependimali cuboidali o colonnari, la cui superficie apicale è ricoperta da microvilli e (parzialmente) ciglia, mentre le superfici laterali sono collegate da complessi di giunzioni intercellulari.
Sostanza bianca del midollo spinale circonda il grigio (vedi Fig. 125) ed è diviso dalle radici anteriori e posteriori in simmetriche retro e corde anteriori.È costituito da fibre nervose che corrono longitudinalmente (principalmente mielinizzate), formando discendenti e ascendenti percorsi (tratti). Questi ultimi sono separati l'uno dall'altro da sottili strati di tessuto connettivo e astrociti, che si trovano anche all'interno dei tratti (Fig. 127). Le vie comprendono due gruppi: propriospinale (effettuano la comunicazione tra le diverse parti del midollo spinale) e le vie sopraspinali (forniscono la comunicazione tra il midollo spinale e le strutture cerebrali - tratti ascendenti e discendenti).
Cervelletto
Cervelletto fa parte del cervello ed è il centro dell'equilibrio, che sostiene
zhaniya tono muscolare e coordinazione dei movimenti. È formato da due emisferi con un gran numero di scanalature e convoluzioni sulla superficie e una parte centrale stretta (verme). materia grigia le forme corteccia cerebellare e noccioli; questi ultimi giacciono nelle sue profondità sostanza bianca.
Corteccia cerebellare caratterizzato da un elevato ordine di localizzazione di neuroni, fibre nervose e cellule gliali di ogni tipo. Si distingue per la ricchezza delle connessioni interneuronali, che assicurano l'elaborazione delle varie informazioni sensoriali che vi entrano. Ci sono tre strati nella corteccia cerebellare (dall'esterno verso l'interno): 1) strato molecolare; 2) strato di cellule di Purkinje (strato di neuroni a forma di pera); 3) strato granulare(Figg. 129 e 130).
strato molecolare contiene un numero relativamente piccolo di piccole cellule, contiene corpi cestino e neuroni stellati. neuroni canestro situato nella parte interna dello strato molecolare. I loro dendriti corti formano legami con fibre parallele nella parte esterna dello strato molecolare, e un lungo assone attraversa il giro, emettendo a determinati intervalli collaterali che scendono ai corpi delle cellule di Purkinje e, ramificandosi, li ricoprono come cesti, formando sinapsi asso-somatiche inibitorie (vedi Fig. 130). neuroni stellati- piccole cellule, i cui corpi si trovano sopra i corpi dei neuroni canestro. I loro dendriti formano connessioni con fibre parallele e le ramificazioni degli assoni formano sinapsi inibitorie sui dendriti delle cellule di Purkinje e possono essere coinvolte nella formazione di un cesto attorno ai loro corpi.
Strato di cellule di Purkinje (strato di neuroni a forma di pera) contiene corpi di cellule di Purkinje che giacciono in una fila, intrecciati con collaterali di assoni di cellule canestro ("cestini").
Cellule di Purkinje (neuroni a forma di pera)- grandi cellule con un corpo a forma di pera contenente organelli ben sviluppati. 2-3 dendriti primari (stelo) partono da esso nello strato molecolare, ramificandosi intensamente con la formazione di dendriti terminali (terminali), raggiungendo la superficie dello strato molecolare (vedi Fig. 130). I dendriti ne contengono numerosi spine- zone di contatto di sinapsi eccitatorie formate da fibre parallele (assoni di neuroni granulari) e sinapsi inibitorie formate da fibre rampicanti. L'assone della cellula di Purkinje parte dalla base del suo corpo, si ricopre di una guaina mielinica, penetra nello strato granulare e penetra nella sostanza bianca, essendo l'unico percorso efferente della sua corteccia.
Strato granulare contiene corpi ravvicinati neuroni granulari, grandi neuroni stellati(cellule del Golgi), così come glomeruli cerebellari- speciali zone di contatto sinaptiche complesse arrotondate tra fibre muschiose, dendriti di neuroni granulari e assoni di grandi neuroni stellati.
Neuroni granulari- i neuroni più numerosi della corteccia cerebellare - piccole cellule con brevi dendriti che assomigliano a un "zampa d'uccello", su cui rosette di fibre muschiose formano numerosi contatti sinaptici nei glomeruli del cervelletto. Gli assoni dei neuroni granulari vengono inviati allo strato molecolare, dove si dividono a forma di T in due rami che corrono paralleli alla lunghezza del giro. (fibre parallele) e formare sinapsi eccitatorie sui dendriti delle cellule di Purkinje, sui neuroni canestro e stellati e sui grandi neuroni stellati.
Grandi neuroni stellati (cellule del Golgi) più grande dei neuroni granulari. I loro assoni all'interno dei glomeruli del cervelletto formano sinapsi inibitorie sui dendriti dei neuroni granulari e lunghi dendriti salgono nello strato molecolare, dove si ramificano e formano connessioni con fibre parallele.
Fibre afferenti della corteccia cerebellare includere briofite e fibre rampicanti(vedi Fig. 130), che penetrano nella corteccia cerebellare dal midollo spinale, dal midollo allungato e dal ponte.
Fibre muscose del cervelletto terminare con le estensioni (prese)- glomeruli del cervelletto, formare contatti sinaptici con i dendriti dei neuroni granulari, sui quali terminano anche gli assoni dei grandi neuroni stellati. I glomeruli del cervelletto non sono completamente circondati all'esterno da processi piatti di astrociti.
Fibre rampicanti del cervelletto penetrano nella corteccia dalla sostanza bianca, passando attraverso lo strato granulare allo strato di cellule di Purkinje e strisciando lungo i corpi e i dendriti di queste cellule, su cui terminano in sinapsi eccitatorie. I rami collaterali delle fibre rampicanti formano sinapsi su altri neuroni di tutti i tipi.
Fibre efferenti della corteccia cerebellare rappresentato dagli assoni delle cellule di Purkinje, che sotto forma di fibre mieliniche vengono inviate alla sostanza bianca e raggiungono i nuclei profondi del cervelletto e il nucleo vestibolare, sui neuroni dei quali formano sinapsi inibitorie (le cellule di Purkinje sono neuroni inibitori).
corteccia cerebraleè l'organizzazione più alta e complessa
ny centro nevralgico, la cui attività assicura la regolazione di varie funzioni dell'organismo e di complesse forme di comportamento. La corteccia è formata da uno strato di materia grigia che ricopre la sostanza bianca, sulla superficie delle convoluzioni e nella profondità dei solchi. La materia grigia contiene neuroni, fibre nervose e cellule neurogliali di ogni tipo. Basato sulle differenze di densità cellulare e struttura (citoarchitettura), percorso della fibra (mieloarchitettura) e le caratteristiche funzionali di varie parti della corteccia al suo interno si distinguono per 52 campi nettamente delimitati.
Neuroni corticali- multipolari, di varie dimensioni e forme, comprendono più di 60 specie, tra le quali si distinguono due tipologie principali - piramidale e non piramidale.
cellule piramidali - tipo di neuroni specifici per la corteccia cerebrale; secondo varie stime, costituiscono il 50-90% di tutti i neuroni corticali. Dal polo apicale del loro corpo a forma di cono (triangolare in sezioni), un lungo dendrite (apicale) ricoperto di spine (Fig. 133) si estende alla superficie della corteccia, dirigendosi verso la placca molecolare della corteccia, dove si dirama . Diversi dendriti laterali (laterali) più corti divergono dalle parti basali e laterali del corpo in profondità nella corteccia e lontano dal corpo del neurone, che, ramificandosi, si diffonde all'interno dello stesso strato in cui si trova il corpo cellulare. Un lungo e sottile assone parte dal centro della superficie basale del corpo, entrando nella sostanza bianca e dando origine ai collaterali. Distinguere cellule piramidali giganti, grandi, intermedie e piccole. La funzione principale delle cellule piramidali è quella di fornire connessioni all'interno della corteccia (cellule intermedie e piccole) e la formazione di vie efferenti (cellule giganti e grandi).
cellule non piramidali situati in quasi tutti gli strati della corteccia, percependo i segnali afferenti in arrivo, e i loro assoni si diffondono all'interno della corteccia stessa, trasmettendo impulsi ai neuroni piramidali. Queste cellule sono molto diverse e sono prevalentemente varietà di cellule stellate. La funzione principale delle cellule non piramidali è l'integrazione dei circuiti neurali all'interno della corteccia.
Citoarchitettura della corteccia cerebrale. I neuroni della corteccia sono disposti in strati non nettamente delimitati (piatti), che sono designati da numeri romani e numerati dall'esterno verso l'interno. Sulle sezioni colorate con ematossilina-eosina, le connessioni tra i neuroni non vengono tracciate, poiché solo
corpi di neuroni e le sezioni iniziali dei loro processi
(Fig. 131).
io - piastra molecolare situato sotto la pia madre; contiene un numero relativamente piccolo di piccoli neuroni orizzontali con lunghi dendriti ramificati che si estendono sul piano orizzontale dal corpo fusiforme. I loro assoni sono coinvolti nella formazione di un plesso tangenziale di fibre di questo strato. Nello strato molecolare sono presenti numerosi dendriti e assoni di cellule di strati più profondi che formano connessioni interneuronali.
II - piastra granulare esterna È formato da numerose piccole cellule piramidali e stellate, i cui dendriti si ramificano e salgono nella piastra molecolare, e gli assoni entrano nella sostanza bianca o formano archi e vanno anche alla piastra molecolare.
III - piastra piramidale esterna caratterizzato dalla predominanza neuroni piramidali, le cui dimensioni aumentano in profondità nello strato da piccolo a grande. I dendriti apicali delle cellule piramidali sono diretti alla piastra molecolare e quelli laterali formano sinapsi con le cellule di questa piastra. Gli assoni di queste cellule terminano all'interno della materia grigia o sono diretti al bianco. Oltre alle cellule piramidali, la lamina contiene una varietà di neuroni non piramidali. La placca svolge funzioni prevalentemente associative, collegando le cellule sia all'interno di un dato emisfero che con l'emisfero opposto.
IV - piastra granulare interna contiene piccola piramidale e cellule stellate. In questa piastra termina la parte principale delle fibre afferenti talamiche. Gli assoni delle cellule di questa lamina formano connessioni con le cellule delle lamine superiori e sottostanti della corteccia.
V - piastra piramidale interna formato grandi neuroni piramidali, e nella regione della corteccia motoria (giro precentrale) - neuroni piramidali giganti(cellule di Betz). I dendriti apicali dei neuroni piramidali raggiungono la placca molecolare, i dendriti laterali si estendono all'interno della stessa placca. Gli assoni di neuroni piramidali giganti e grandi sono proiettati ai nuclei del cervello e del midollo spinale, i più lunghi nella composizione delle vie piramidali raggiungono i segmenti caudali del midollo spinale.
VI - piatto multiforme formato da neuroni di varie forme, e la sua
le aree esterne contengono cellule più grandi, mentre le aree interne contengono cellule più piccole e scarsamente posizionate. Gli assoni di questi neuroni entrano nella sostanza bianca come parte delle vie efferenti e i dendriti penetrano nella plasticità molecolare.
Mieloarchitettura della corteccia cerebrale. Le fibre nervose della corteccia cerebrale comprendono tre gruppi: 1) afferente; 2) associativo e commissario; 3) efferente.
Fibre afferenti arrivano alla corteccia dalle parti inferiori del cervello sotto forma di fasci nella composizione strisce verticali- travi radiali (vedi Fig. 132).
Fibre di associazione e commissurali - fibre intracorticali che collegano diverse aree della corteccia all'interno di uno o in diversi emisferi, rispettivamente. Queste fibre formano fasci (strisce) che corrono parallele alla superficie della corteccia nella piastra I (piatto tangenziale), nella tavola II (piastra disfibrosa, o striscia di Bechterew), nella tavola IV (striscia di lamiera granulare esterna, o fascia esterna di Bayarzhe) e nella tavola V (striscia di lamina granulare interna, o la fascia interna di Bayarzhe) - vedi fig. 132. Gli ultimi due sistemi sono plessi formati dalle sezioni terminali di fibre afferenti.
Fibre efferenti collegare la corteccia con le formazioni sottocorticali. Queste fibre corrono verso il basso come parte dei raggi radiali.
Tipi di struttura della corteccia cerebrale.
In alcune aree della corteccia associate allo svolgimento di varie funzioni, predomina lo sviluppo di alcuni strati di essa, sulla base del quale si distinguono agranulare e tipi granulari di corteccia.
Tipo di corteccia agranulare caratteristico dei suoi centri motori e si distingue per il maggior sviluppo delle placche III, V e VI della corteccia con un debole sviluppo delle placche II e IV (granulari). Tali aree della corteccia fungono da fonti di percorsi discendenti.
Tipo granulare di corteccia caratteristica delle aree di localizzazione dei centri corticali sensibili. Si distingue per un debole sviluppo di strati contenenti cellule piramidali, con una significativa gravità delle placche granulari (II e IV).
Sostanza bianca del cervello rappresentato da fasci di fibre nervose che salgono alla materia grigia della corteccia dal tronco cerebrale e scendono al tronco cerebrale dai centri corticali della materia grigia.
ORGANI DEL SISTEMA NERVOSO
Organi del sistema nervoso periferico
Riso. 114. Nervo (tronco nervoso). sezione trasversale
Colore: osmirovania
1 - fibre nervose; 2 - endoneurio; 3 - perineurio; 4 - epinevrio: 4.1 - tessuto adiposo, 4.2 - vaso sanguigno

Riso. 115. Sezione di un nervo (tronco nervoso)
Colore: osmirovania
1 - fibra mielinica: 1.1 - processo neuronale, 1.2 - guaina mielinica;
2- fibra amielinica; 3 - endoneurio; 4 - perineurio

Riso. 116. Tronco nervoso (nervo). sezione trasversale
Colorazione: ematossilina-eosina
1 - fibre nervose; 2 - endoneurium: 2.1 - vaso sanguigno; 3 - perineurio; 4 - epinevrio: 4.1 - cellule adipose, 4.2 - vasi sanguigni

Riso. 117. Sezione del tronco nervoso (nervo)
Colorazione: ematossilina-eosina
1 - fibra mielinica: 1.1 - processo neuronale, 1.2 - guaina mielinica, 1.3 - nucleo dei neurolemmociti; 2 - fibra amielinica; 3 - endoneurium: 3.1 - vaso sanguigno; 4 - perineurio; 5 - epinevrio

Riso. 118. Sezione del tronco nervoso (nervo)
1 - fibra mielinica: 1.1 - processo neuronale, 1.2 - guaina mielinica; 2 - fibra amielinica; 3 - endoneurium: 3.1 - vaso sanguigno; 4 - perineurio

Riso. 119. Arco riflesso somatico
1.Collegamento del recettore formato neuroni afferenti (sensoriali) pseudounipolari, i cui corpi (1.1) si trovano nei nodi sensoriali del nervo spinale (1.2). I processi periferici (1.3) di queste cellule formano le terminazioni nervose sensoriali (1.4) nella pelle o nel muscolo scheletrico. I processi centrali (1.5) entrano nel midollo spinale come parte di radici posteriori(1.6) e vengono inviati a corna posteriori di materia grigia formare sinapsi sui corpi e dendriti dei neuroni intercalari (archi riflessi a tre neuroni, A), o passare nelle corna anteriori ai motoneuroni (archi riflessi a due neuroni, B).
2.Collegamento associativo presentato (2.1), i cui dendriti e corpi giacciono nelle corna posteriori. I loro assoni (2.2) vengono inviati a corna anteriori, trasmettere impulsi nervosi ai corpi e ai dendriti dei neuroni effettori.
3.Collegamento efferente formato motoneuroni multipolari(3.1). I corpi e i dendriti di questi neuroni si trovano nelle corna anteriori, formando i nuclei motori. Gli assoni (3.2) dei motoneuroni lasciano il midollo spinale come parte di radici anteriori(3.3) e inoltre come parte del nervo misto (4) sono inviati al muscolo scheletrico, dove i rami dell'assone formano sinapsi neuromuscolari (3.4)

Riso. 120. Arco riflesso autonomo (vegetativo).
1.Collegamento del recettore formato neurone pseudounipolare afferente (sensoriale). mi, i cui corpi (1.1) giacciono nei nodi sensoriali del nervo spinale (1.2). I processi periferici (1.3) di queste cellule formano le terminazioni nervose sensoriali (1.4) nei tessuti degli organi interni. I processi centrali (1.5) entrano nel midollo spinale come parte di dietro di loro mozziconi(1.6) e vengono inviati a corna laterali di materia grigia formare sinapsi sui corpi e sui dendriti degli interneuroni.
2.Collegamento associativo presentato interneuroni multipolari(2.1), i cui dendriti e corpi si trovano nelle corna laterali del midollo spinale. Gli assoni di questi neuroni sono fibre pregangliari (2.2). Lasciano il midollo spinale come parte di radici anteriori(2.3), dirigendosi verso uno dei gangli autonomi, dove terminano sui corpi e sui dendriti dei loro neuroni.
3.Collegamento efferente formato multipolare o neuroni bipolari, i cui corpi (3.1) giacciono in gangli autonomi (3.2). Gli assoni di queste cellule sono fibre postgangliari (3.3). Come parte dei tronchi nervosi e dei loro rami, vengono inviati alle cellule degli organi di lavoro: muscoli lisci, ghiandole, cuore, che formano terminazioni su di essi (3.4). Nei gangli vegetativi, oltre ai neuroni efferenti "ad assoni lunghi" - cellule di Dogel tipo I (DI), ci sono neuroni afferenti "ugualmente escrescenze" - cellule di Dogel di tipo II (DII), che fanno parte degli archi riflessi locali come un collegamento di recettore e gabbie associative di tipo III Dogelya (DIII) - piccoli neuroni intercalari

Riso. 121. Ganglio sensoriale del nervo spinale
Colorazione: ematossilina-eosina
1 - colonna vertebrale posteriore; 2 - ganglio sensibile del nervo spinale: 2.1 - capsula del tessuto connettivo, 2.2 - corpi di neuroni sensoriali pseudo-unipolari, 2.3 - fibre nervose; 3 - dorso anteriore; 4 - nervo spinale

Riso. 122. Neurone pseudounipolare del ganglio sensoriale del nervo spinale e del suo microambiente tissutale
Colorazione: ematossilina-eosina
1 - corpo di un neurone sensibile pseudounipolare: 1.1 - nucleo, 1.2 - citoplasma; 2 - cellule gliali satelliti; 3 - capsula di tessuto connettivo attorno al corpo del neurone

Riso. 123. Ganglio autonomo (vegetativo) dal plesso solare
1 - fibre nervose pregangliari; 2 - ganglio autonomo: 2.1 - capsula del tessuto connettivo, 2.2 - corpi di neuroni autonomi multipolari, 2.3 - fibre nervose, 2.4 - vasi sanguigni; 3 - fibre postgangliari

Riso. 124. Neurone multipolare del ganglio autonomo e del suo microambiente tissutale
Colorazione: ematossilina di ferro
1 - corpo di un neurone multipolare: 1.1 - nucleo, 1.2 - citoplasma; 2 - l'inizio dei processi; 3 - gliociti; 4 - guaina del tessuto connettivo
Organi del sistema nervoso centrale

Riso. 125. Midollo spinale (sezione trasversale)
Colore: nitrato d'argento
1 - materia grigia: 1.1 - corno anteriore (ventrale), 1.2 - corno posteriore (dorsale), 1.3 - corno laterale (laterale); 2 - aderenze grigie anteriori e posteriori: 2.1 - canale centrale; 3 - fessura mediana anteriore; 4 - solco mediano posteriore; 5 - sostanza bianca (tratti): 5.1 - cordone dorsale, 5.2 - cordone laterale, 5.3 - cordone ventrale; 6 - guscio molle del midollo spinale

Riso. 126. Midollo spinale.
Area di materia grigia (corna anteriori)
Colorazione: ematossilina-eosina
1- corpi di motoneuroni multipolari;
2- gliociti; 3 - neuropillo; 4 - vasi sanguigni

Riso. 127. Midollo spinale. Area di sostanza bianca
Colorazione: ematossilina-eosina
1 - fibre nervose mielinizzate; 2 - nuclei di oligodendrociti; 3 - astrociti; 4 - vaso sanguigno

Riso. 128. Midollo spinale. Canale centrale
Colorazione: ematossilina-eosina
1 - ependimociti: 1.1 - ciglia; 2 - vaso sanguigno

Riso. 129. Cervelletto. Abbaio
(fetta perpendicolare al corso delle convoluzioni)
Colorazione: ematossilina-eosina
1 - guscio molle del cervello; 2 - materia grigia (corteccia): 2.1 - strato molecolare, 2.2 - strato di cellule di Purkinje (neuroni a forma di pera), 2.3 - strato granulare; 3 - sostanza bianca

Riso. 130. Cervelletto. Trama di corteccia
Colore: nitrato d'argento
1 - strato molecolare: 1.1 - dendriti delle cellule di Purkinje, 1.2 - fibre afferenti (rampicanti), 1.3 - neuroni dello strato molecolare; 2 - strato di cellule di Purkinje (neuroni a forma di piri): 2.1 - corpi di neuroni a forma di pera (cellule di Purkinje), 2.2 - "cestini" formati da collaterali di assoni di neuroni canestro; 3 - strato granulare: 3.1 - corpi di neuroni granulari, 3.2 - assoni di cellule di Purkinje; 4 - sostanza bianca

Riso. 131. Emisfero cerebrale. Abbaio. Citoarchitettura
Colorazione: ematossilina-eosina
1 - guscio molle del cervello; 2 - materia grigia: le placche (strati) della corteccia sono indicate con numeri romani: I - placca molecolare, II - placca granulare esterna, III - placca piramidale esterna, IV - placca granulare interna, V - placca piramidale interna, VI - multiforme piatto; 3 - sostanza bianca
Riso. 132. Emisfero cerebrale. Abbaio.
Mieloarchitettura
(schema)
1 - piatto tangenziale; 2 - placca disfibrosa (striscia di Bekhterev); 3 - raggi radiali; 4 - striscia della piastra granulare esterna (striscia esterna di Bayarzhe); 5 - striscia di piastra granulare interna (striscia interna di Bayarzhe)

Riso. 133. Grande neurone piramidale dell'emisfero cerebrale
Colore: nitrato d'argento
1 - grande neurone piramidale: 1.1 - corpo del neurone (pericarione), 1.2 - dendriti, 1.3 - assone;
2- gliociti; 3 - neuropillo
Particolari
gangli rappresentare cluster di neuroni multipolari (un assone e diversi dendriti).(da poche cellule a decine di migliaia). I gangli extraorganici (simpatici) hanno una capsula di tessuto connettivo ben definita, come continuazione del perineurio. I gangli parasimpatici si trovano solitamente nei plessi nervosi intramurali. I gangli dei plessi intramurali, come altri nodi autonomici, contengono neuroni autonomi di archi riflessi locali. I neuroni multipolari con un diametro di 20-35 μm si trovano in modo diffuso, ogni neurone è circondato da gliociti gangliari.
Inoltre, descritto neuroendocrini, chemocettori, bipolari e, in alcuni vertebrati, neuroni unipolari. Nei gangli simpatici sono presenti piccole cellule intensamente fluorescenti (cellule MYF) con processi brevi e un gran numero di vescicole granulari nel citoplasma. Secernono catecolamine e hanno un effetto inibitorio sulla trasmissione degli impulsi dalle fibre nervose pregangliari al neurone simpatico efferente. Queste cellule sono chiamate interneuroni.
Tra i maggiori neuroni multipolari vegetativo gangli distinguere: motore (le cellule di Dogel del 1o tipo), sensibile (le cellule di Dogel del II-esimo tipo) e associativo (le cellule di Dogel del III-esimo tipo). I motoneuroni hanno dendriti corti con estensioni lamellari ("cuscinetti ricettivi"). L'assone di queste cellule è molto lungo, va oltre il ganglio come parte delle sottili fibre nervose non mielinizzate postgangliari e termina sui miociti lisci degli organi interni. Le cellule del 1° tipo sono chiamate neuroni ad assone lungo. I neuroni del II-esimo tipo sono cellule nervose equidistanti. 2-4 processi partono dal loro corpo, tra i quali è difficile distinguere un assone. Senza ramificazioni, i processi si allontanano dal corpo del neurone. I loro dendriti hanno terminazioni nervose sensibili e l'assone termina sui corpi dei motoneuroni nei gangli vicini. Le cellule di tipo II sono neuroni sensibili degli archi riflessi autonomici locali. Le cellule Dogel di tipo III hanno una forma corporea simile ai neuroni autonomi di tipo II, ma i loro dendriti non si estendono oltre il ganglio e il neurite va ad altri gangli. Molti ricercatori considerano queste cellule come varietà di neuroni sensibili.
Pertanto, nei gangli autonomici periferici ci sono archi riflessi locali costituiti da neuroni autonomici sensoriali, motori e possibilmente associativi.
I gangli autonomi intramurali nella parete del tubo digerente si distinguono per il fatto che, oltre ai neuroni colinergici motori, contengono neuroni inibitori. Sono rappresentati da cellule nervose adrenergiche e purinergiche. In quest'ultimo, il mediatore è un nucleotide purinico. Nei gangli autonomici intramurali ci sono anche neuroni peptidergici che secernono il peptide vasointestinale, la somatostatina e una serie di altri peptidi, con l'aiuto dei quali viene eseguita la regolazione neuroendocrina e la modulazione dell'attività dei tessuti e degli organi dell'apparato digerente.
Acetilcolina- recettori nicotinici (blocco curativo, esametonio), muscarinici (blocco dell'atropina). Attivazione del recettore → Generazione EPSP. EPSP rapido (N-colinergico)→apertura dei canali ionici. EPSP lento (M-colinergico) → soppressione della corrente M a causa di un aumento della conduttività K.
Neuropeptidi- agiscono come neuromodulatori.

encefaline, sostanza P, luliberina, neurotensina, somatostatina - sintomo. gangli (+Ach)
catecolamine(NA, dopamina) sono neurotrasmettitori a piccole cellule con fluorescenza intensa.
Neuropeptide Y, somatostatina - sintomo. postgangliare.
Cellule postgangliari simpatiche: NA, ATP, neuropeptide U.
α1→inosotolo trifosfato, diacilglicerolo. α2→Attivazione della proteina G, ↓cAMP.
β→proteina G→AC→cAMP
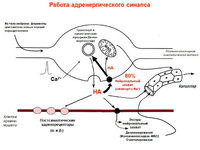
Eccezioni: mediatore Ach, recettori muscarinici.
Parasimpatico. postgangliari: Ach, VIP, NO, somatostatina, ATP, peptidi oppioidi.
M1 (alta affinità per la pirenzepina) - aumento della secrezione acida da parte delle cellule delle ghiandole dello stomaco, M2 (bassa) - rallenta il cuore. ritmo, secrezione delle ghiandole lacrimali e salivari.
Azione varia:
-Specifico sec. mediatori: M2 può agire IP3, oppure può indurre AC, riducendo cAMP.
- Azione sui canali K e Ca
-NO → guanilato ciclasi → cGMP → protein chinasi cGMP-dipendente → il rilassamento della muscolatura liscia si forma sull'endotelio.