Ganglios linfáticos: número, estructura, aumento (linfadenopatía). Senos linfáticos y migración celular
Página 6 de 6
LOS GANGLIOS LINFÁTICOS
Los ganglios linfáticos (hodi linfatici) son los órganos más numerosos sistema inmunitario, sirven como filtros biológicos para la linfa (líquido tisular) que fluye a través de ellos. Yacen en el camino vasos linfáticos de órganos y tejidos a conductos linfaticos y troncos linfáticos. Los ganglios linfáticos generalmente se ubican en grupos. Puede haber dos o más, ya veces varias docenas de nodos en un grupo. Por ejemplo, el grupo de ganglios mesentéricos superiores tiene 66-404, axilar - 12-45, inguinal superficial - 4-20 ganglios.
Cada ganglio linfático(nodus linfaticus) está cubierto en el exterior con una cápsula de tejido conectivo, desde la cual se extienden ramas delgadas dentro del órgano: barras transversales, trabéculas capsulares(trabéculas). En el lugar donde los vasos linfáticos eferentes salen del ganglio linfático, el ganglio tiene una ligera depresión: puertas(hilio). En la región de la puerta, la cápsula se engrosa bastante, formando un engrosamiento portal (hiliar), que sobresale más o menos profundamente en el nódulo. Las trabéculas portales se extienden desde el engrosamiento portal hacia el parénquima del nódulo. El más largo de ellos se conecta con las trabéculas capsulares.
En los ganglios linfáticos, a los que fluye la linfa desde las extremidades (inguinal, axilar) y que también se denominan omáticos, más a menudo una puerta, en visceral (mesentérica, traqueobronquial), hasta 3-4 puertas. A través de la puerta, las arterias, los nervios ingresan al ganglio linfático, las venas y los vasos linfáticos eferentes salen.
Dentro del ganglio linfático, entre las trabéculas, se encuentra el estroma reticular. Está representado por fibras reticulares y células reticulares que forman una red tridimensional con bucles de varios tamaños y formas.
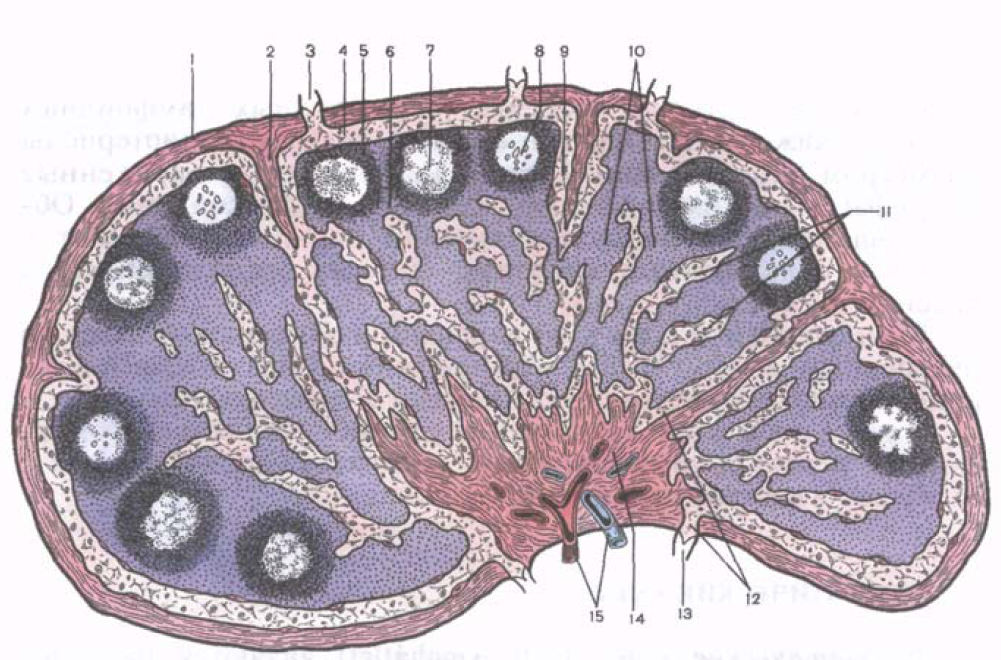
Arroz. 39. Estructura microscópica ganglio linfático.
1 - cápsula; 2 - trabécula; 3 - trayendo el vaso linfático; 4 - seno linfático subcapsular; 5 - sustancia cortical; 6 - zona paracortical (dependiente del timo); 7 - nódulo linfoide; 8 - el centro de reproducción del nódulo linfoide; 9 - seno linfático cortical; 10 - hebras carnosas; 11 - senos cerebrales; 12 - seno portal; 13 - vaso linfático eferente; 14 - engrosamiento del portal; 15 - vasos sanguíneos.
Los elementos celulares del tejido linfoide se encuentran en las asas del estroma reticular.
El parénquima del ganglio linfático se divide en cortical y medular (Fig. 39). La corteza es más oscura en las secciones histológicas teñidas debido a la densa densidad de elementos celulares, está más cerca de la cápsula y ocupa las partes periféricas del nódulo. Encendedor médula(médula) se encuentra más cerca de la puerta del nodo y ocupa su parte central. En la sustancia cortical hay formaciones redondeadas con un diámetro de 0,5-1,0 mm - nódulos linfoides(nódulos linfoides). Hay nódulos linfoides sin centro de crianza y con Centro de cría(centro germinativo, centrum germinale).
El tejido linfoide difuso se encuentra alrededor de los nódulos linfoides. En él se distingue una meseta cortical, que incluye áreas de tejido linfoide entre los nódulos, la zona internodular. La composición de la meseta cortical también incluye tejido ubicado fuera de los nódulos linfoides, entre estos y la cápsula. Hacia adentro de los nódulos, directamente en el borde con la médula, se destaca una tira de tejido linfoide: la sustancia pericortical, o zona dependiente del timo (paracortical)(paracortex, s.zona thymodedependens), que contiene predominantemente linfocitos T. En esta zona hay vénulas poscapilares revestidas con un endotelio cúbico, a través de cuyas paredes migran los linfocitos al torrente sanguíneo.
El parénquima de la médula está representado por hebras de tejido linfoide: hebras pulposas(cuerdas medulares). se extienden desde departamentos internos sustancia cortical a la puerta del ganglio linfático y junto con los nódulos linfoides forman una zona B-dependiente. Las hebras de pulpa están conectadas entre sí, lo que da como resultado tejidos complejos.
El parénquima del ganglio linfático está impregnado de una densa red de canales estrechos: senos linfáticos(sinus linfatici), a través del cual la linfa que ingresa al ganglio fluye desde seno subcapsular (marginal)(seno subcapsular) al seno porta. A lo largo de las trabéculas capsulares se encuentran senos corticales(senos corticales) y médula(seno medular). Estos últimos alcanzan la puerta del ganglio linfático (engrosamiento del portal) y fluyen hacia el seno del portal que se encuentra aquí. En la luz de los senos paranasales hay una red de bucles pequeños formada por fibras y células reticulares, en cuyos bucles pueden atascarse partículas extrañas, células muertas y tumorales.
Desarrollo y caracteristicas de la edad ganglios linfáticos Los ganglios linfáticos y su estroma se desarrollan a partir del mesénquima cerca de los plexos emergentes de vasos sanguíneos y linfáticos, a partir de la semana 5-6 de la vida del embrión. Los marcadores de los ganglios linfáticos en diferentes áreas del cuerpo humano se forman en diferentes periodos antes del nacimiento e incluso después de él.
En el proceso de desarrollo de los ganglios, la luz del vaso linfático se convierte en un seno subcapsular (marginal). Los senos intermedios se desarrollan sobre la base de un plexo linfático ramificado, entre cuyos vasos crecen hebras de tejido conectivo embrionario. Las células de la serie linfoide se infunden aquí. A partir de la semana 19, en algunos ganglios linfáticos se puede ver el borde delimitado entre la corteza y la médula. Los nódulos linfoides en los ganglios linfáticos comienzan a formarse ya en el período prenatal. Los centros de reproducción en los nódulos linfoides aparecen poco antes y poco después del nacimiento. Los principales procesos de conformación relacionados con la edad en los ganglios linfáticos terminan entre los 10 y los 12 años.
Cambios relacionados con la edad en ganglios linfáticos(disminución de la cantidad de linfoides y proliferación de tejido adiposo) se observan ya en la adolescencia. El tejido conectivo crece en el estroma y el parénquima de los ganglios, aparecen grupos de células grasas. Al mismo tiempo, disminuye el número de ganglios linfáticos en grupos regionales. Muchos ganglios linfáticos pequeños son reemplazados por completo por tejido conectivo y dejan de existir como órganos del sistema inmunitario. Los ganglios linfáticos cercanos, a menudo de tamaño mediano, se fusionan entre sí y forman ganglios más grandes de forma segmentaria o similar a una cinta.
Vasos y nervios de los ganglios linfáticos. Cada ganglio linfático incluye de 1 a 2 a 10 ramas arteriales pequeñas de las arterias más cercanas. Las vénulas formadas a partir de los capilares se fusionan en venas, van a las puertas del órgano y salen del nódulo junto con los vasos linfáticos eferentes.
Vegetativo fibras nerviosas Los ganglios linfáticos se obtienen de los plexos ubicados junto a las arterias, así como de los troncos nerviosos que pasan cerca de los ganglios.
Preguntas para la autorrepetición y el autocontrol
1. ¿Qué órganos son los órganos centrales del sistema inmunitario, cuáles son los periféricos y por qué?
2. ¿Qué características morfológicas son típicas de los órganos centrales del sistema inmunitario y cuáles de los periféricos?
3. ¿Recuerdas en qué elementos estructurales consisten la médula ósea y el timo?
4. ¿Qué es común en la estructura de los órganos periféricos del sistema inmunitario?
5. ¿En las paredes de qué órganos se localizan los nódulos linfoides? ¿Cuál es su estructura?
6. ¿Qué elementos estructurales del tejido linfoide están incluidos en la pulpa blanca del bazo?
7. ¿Qué formaciones hay en los ganglios linfáticos, a través de los cuales fluye la linfa desde los vasos linfáticos aferentes a los eferentes?
Estado de Cheliábinsk academia medica
Departamento de Histología, Citología y Embriología
Conferencia
Órganos periféricos de hematopoyesis e inmunogénesis.
Lista de diapositivas.
1. Tejido reticular del ganglio linfático (335).
2. Tejido reticular del bazo (esquema) (336).
3. Ganglio linfático (esquema) (339).
4. Ganglio linfático. Cordones carnosos (diagrama) (338).
5. Ganglio linfático. Senos paranasales (340).
6. Bazo (diagrama) (341).
7. Bazo (342).
8. Bazo. folículo linfoide. arteria central (343)
9. Senos cerebrales de un ganglio linfático no estimulado (390).
10. Senos cerebrales del ganglio linfático estimulado (391).
11. Fibras reticulares (esquema) (337).
12. Células plasmáticas del bazo de ratón (373).
13. Macrófagos, linfocitos, blastocitos en un ganglio linfático (371).
14. Célula dendrítica y linfocito (376).
15. Linfoblastos y linfocitos en el bazo (369).
16. Folículos linfoides de la placa de Peyre (355).
17. Zona B de un ganglio linfático de un ratón inmunizado con una fuerte transformación blástica y vénulas poscapilares activas (386).
Plan.
1. características generalesórganos periféricos de hematopoyesis y defensa inmune.
2. Histofisiología del ganglio linfático: fuente de desarrollo, estructura, características de la cortical y la médula, zonas T y B, funciones.
3. Características del tejido linfoide.
4. Histofisiología del bazo: fuente de desarrollo, estructura, características de la corteza y la médula, zonas T y B, funciones.
5. Linfopoyesis T y B.
Propósito de la conferencia:
1. Familiarizarse con las características morfológicas y funcionales de los órganos periféricos de hematopoyesis e inmunodefensa.
2. Dar el concepto de tejido linfoide.
3. Describir en detalle las características de la linfopoyesis T y B.
Los órganos hematopoyéticos periféricos incluyen ganglios linfáticos, bazo, amígdalas, proceso apendicular y folículos linfoides en la pared. tubo digestivo, vías respiratorias y sistema urinario. En los órganos periféricos de la hematopoyesis, hay una reunión de células inmunocompetentes con antígenos. Después de eso, se activan las respuestas inmunitarias, que se basan en la diferenciación de linfocitos dependiente del antígeno, lo que da como resultado la formación de células efectoras que inactivan el antígeno, incluidos los asesinos de T citotóxicos y las células plasmáticas que producen anticuerpos.
Los ganglios linfáticos.
En un adulto, hay hasta 1000 ganglios linfáticos, cuyo tamaño varía desde el tamaño de una cabeza de alfiler hasta un pequeño grano de frijol (promedio de 1 cm).
El ganglio linfático tiene forma de frijol y está ubicado a lo largo del curso de los vasos linfáticos. Desde la superficie convexa, ingresan los vasos linfáticos aferentes. La superficie cóncava del ganglio linfático se llama puerta. En la región de la puerta, una arteria y nervios ingresan al ganglio linfático y salen una vena y un vaso linfático. Desde la superficie, el ganglio linfático está cubierto con una cápsula que consiste en tejido conectivo denso, en cuyas capas profundas hay células de músculo liso que promueven la promoción de la linfa. El contenido de células musculares lisas y fibras elásticas en las cápsulas de los ganglios linfáticos no es el mismo. Por lo tanto, los ganglios linfáticos inguinales y mesentéricos en humanos son ricos en células musculares. Al mismo tiempo, los ganglios linfáticos seniles se caracterizan, por regla general, por un mayor contenido de fibras elásticas. Alrededor de la cápsula de tejido conectivo se encuentra una poderosa capa de tejido adiposo, que rodea constantemente el ganglio linfático.
Las trabéculas fibrosas se extienden desde la superficie interna de la cápsula y se anastomosan entre sí en las partes centrales de los ganglios linfáticos. El estroma del ganglio linfático está representado por tejido conjuntivo laxo y sin forma y tejido reticular. Aparecen grupos de linfocitos en el tejido reticular del ganglio linfático. El tejido reticular infiltrado con linfocitos se denomina tejido linfoide. A partir de la cápsula del ganglio linfático, el tejido linfoide forma racimos esféricos, que se denominan nódulos secundarios o folículos linfoides. Desde los folículos linfoides, las hebras se extienden hacia el interior del nódulo, llamadas cordones medulares (cordones cerebrales). Los cordones pulmonares se anastomosan repetidamente entre sí. La presencia de nódulos secundarios (folículos linfoides) situados a lo largo de la periferia, y cordones pulposos (cordones cerebrales) que ocupan la parte central del ganglio, permiten aislar la cortical y la médula del ganglio linfático. Entre la cortical y la médula se encuentra la zona paracortical.
La sustancia cortical se encuentra a lo largo de la periferia del nódulo y está representada por folículos linfoides, en los que la parte central tiene un color claro y la parte periférica es más oscura. La parte central se llama centro de luz, que contiene principalmente linfocitos grandes (inmaduros). Debido a que en esta zona se encuentran numerosas mitosis, se le denominó centro de reproducción. Debido al hecho de que bajo la influencia del antígeno en esta zona, los procesos de proliferación de linfocitos se desarrollan de manera rápida y bastante intensa, se denomina centro reactivo. Los centros de luz (centros de reproducción) no se encuentran en todos los folículos, y sus tamaños son muy variables. Entonces, en animales mantenidos en condiciones asépticas, no hay focos de luz. La parte periférica, teñida de oscuro, del folículo linfoide se denomina zona del manto, en la que predominan los linfocitos pequeños. Los folículos linfoides no son formaciones permanentes: pueden aparecer y desaparecer nuevamente.
Los folículos linfoides y los cordones pulposos están rodeados por senos de ganglios linfáticos, que son espacios en forma de hendidura llenos de tejido reticular. La linfa fluye a través de los senos paranasales. Hay varios tipos de senos paranasales, incluidos el seno marginal, el seno cortical intermedio, el seno cerebral y el seno porta. El seno marginal (subcapsular) está limitado por la cápsula de tejido conectivo del ganglio linfático y la superficie del folículo linfoide. El seno cortical intermedio está delimitado por la superficie de las trabéculas y la superficie lateral del folículo linfoide. Los senos cerebrales se encuentran entre los cordones pulposos ramificados. El seno portal (seno terminal) se encuentra en la región de la puerta del ganglio linfático. La pared de todos los senos paranasales del ganglio linfático está revestida con células reticulares modificadas especiales: células costeras. Las células costeras adquieren características de células endoteliales con alta actividad fagocítica. Existen numerosos poros entre las células costeras, que establecen amplias conexiones entre las luces de los senos paranasales y el tejido linfoide de los cordones pulposos y los folículos linfoides. Gracias a las células costeras, la linfa que fluye a través de los senos paranasales se filtra y se limpia de compuestos extraños.
En el ganglio linfático, se distinguen las zonas T y B. La zona B está representada por folículos linfoides y cordones pulposos. En esta zona, la diferenciación dependiente de antígeno de los linfocitos B ocurre bajo la influencia de un microambiente específico, que incluye células reticulares, células dendríticas del primer tipo, macrófagos y una pequeña cantidad de linfocitos T. La zona T está representada por el tejido linfoide de la zona paracortical. En esta zona, la diferenciación dependiente de antígeno de los linfocitos T ocurre bajo la influencia de un microambiente específico, que incluye células reticulares, células dendríticas del segundo tipo (células interdigitadas), macrófagos, así como una pequeña cantidad de linfocitos B y células plasmáticas. .
En la zona paracortical, hay secciones especializadas del torrente sanguíneo: "vénulas poscapilares", cuya pared está revestida con células endoteliales cúbicas o prismáticas. Estas células endoteliales tienen un retículo endoplásmico bien desarrollado, vesículas citoplasmáticas y microvellosidades. Las vénulas poscapilares son el sitio de invasión de los linfocitos T y B en el ganglio linfático. Los linfocitos T y B se adhieren inicialmente a las células endoteliales y luego pasan entre ellas, adquiriendo polaridad citoplasmática. Después de penetrar en el tejido linfoide, los linfocitos pueblan las zonas T y B donde tiene lugar su diferenciación dependiente de antígeno.
Los ganglios linfáticos están ricamente inervados. Las fibras nerviosas amielínicas y no mielinizadas ingresan al ganglio linfático junto con la arteria.
Funciones del ganglio linfático.
1. Función linfopoyética (hematopoyética). La linfa que fluye a través de los senos paranasales del ganglio linfático se enriquece con linfocitos T y B maduros, dejando el tejido linfoide a través de los poros entre las células costeras.
2. Función inmunológica. Debido a los linfocitos T y B formados en los ganglios linfáticos, estos últimos están involucrados en la regulación de la inmunidad celular y humoral.
3. Función de barrera (protectora). La linfa que fluye a través de los senos paranasales se elimina de compuestos extraños debido a la actividad fagocítica de las células costeras.
4. Función de depósito. Normalmente, una cierta cantidad de linfa permanece en el ganglio linfático y se desconecta del flujo linfático. Si es necesario, vuelve a entrar en la circulación linfática.
5. Función de intercambio. Los ganglios linfáticos toman Participación activa en el metabolismo, incluyendo proteínas, grasas, carbohidratos y otros compuestos. Los nutrientes en la linfa pueden ser capturados por los macrófagos y degradados por las enzimas lisosomales.
Desarrollo. Los ganglios linfáticos se desarrollan en la semana 8-10 de la embriogénesis a partir de acumulaciones de células mesenquimales cerca de los vasos sanguíneos y a lo largo de los vasos linfáticos. A lo largo de la periferia de estas acumulaciones, se forma una cápsula de tejido conjuntivo a partir del mesénquima. Entre el mesénquima del rudimento del nódulo y la cápsula, se forma un espacio: el seno subcapsular, del cual parten los senos perinodales corticales, que separan los nódulos entre sí; los senos cerebrales parten de los senos perinodulares. Las trabéculas se extienden desde la cápsula de tejido conjuntivo profundamente hacia el rudimento mesenquimatoso del nódulo. Las células mesenquimatosas del rudimento del ganglio linfático se diferencian en su estroma reticular. Pronto, las células madre invaden los rudimentos de los ganglios linfáticos y comienza la mielopoyesis, que dura un período de tiempo relativamente corto. En la semana 16, los linfocitos B invaden el centro de los ganglios linfáticos, unos días después (los linfocitos B) también invaden sus partes periféricas, y los linfocitos T son los últimos en invadir. A partir de este momento comienza la linfopoyesis en los ganglios linfáticos, que continúa antes y después del nacimiento. Hacia la semana 20 de la embriogénesis, los ganglios linfáticos adquieren las características de los ganglios linfáticos finales (definitivos).
Estructura. Los ganglios linfáticos se encuentran a lo largo de los vasos sanguíneos y linfáticos, tienen forma ovalada o en forma de frijol. En el exterior, los ganglios linfáticos están cubiertos por una cápsula de tejido conectivo, desde la cual se extienden las trabéculas. La cápsula y las trabéculas incluyen colágeno y fibras elásticas y miocitos lisos. En la superficie cóncava de los ganglios linfáticos hay puertas. La arteria y los nervios entran por la puerta, las venas y el vaso linfático eferente salen. Los vasos linfáticos aferentes entran por el lado convexo opuesto. El estroma de los ganglios linfáticos es tejido reticular, que consta de células reticulares y fibras reticulares. El diámetro de los ganglios linfáticos es de 0,5 a 1 cm.
En la periferia de los ganglios linfáticos hay una sustancia cortical más oscura, representada por los ganglios linfáticos, y en el centro, una médula más clara, representada por los cordones medulares. Los ganglios linfáticos tienen una zona cortical (corteza), una médula y una zona paracortical ubicada entre la zona cortical (nódulos linfáticos) y las hebras de la médula.
zona cortical representado por nódulos linfáticos (nódulo linfático), cuyo diámetro varía de 0,5 a 1 mm. El estroma de los nódulos linfáticos está representado por tejido reticular, predominantemente fibras reticulares ubicadas circularmente. Los nódulos linfáticos contienen macrófagos libres, células dendríticas, linfocitos B y linfoblastos B. En el centro de los ganglios linfáticos se encuentra luz centro (centrum lucidum). Este centro también se denomina centro germinal, ya que aquí se multiplican los linfoblastos B. así como el centro reactivo, porque aquí tiene lugar la reacción entre los macrófagos y los antígenos. En otras palabras, en los centros de luz hay una activación de linfocitos por antígenos, reproducción de linfoblastos (inmunoblastos, plasmablastos) y fagocitosis de antígenos y células linfoides.
macrófagos Los nódulos linfáticos realizan una función fagocítica y procesan antígenos desde un estado corpuscular a un estado molecular, acumulando moléculas de antígeno en una cantidad tal que puede causar la diferenciación dependiente de antígeno de los linfocitos B.
Células dendríticas Los nódulos linfáticos son macrófagos que han perdido la capacidad de fagocitar. En las células dendríticas hay procesos, orgánulos poco desarrollados de importancia general. El citoplasma de estas células está débilmente teñido, en su superficie hay receptores para inmunoglobulinas. Las inmunoglobulinas se unen a estos receptores y los antígenos se unen a los extremos libres de las inmunoglobulinas. Los antígenos de las células dendríticas, junto con los antígenos de los macrófagos y con la participación de los ayudantes T, estimulan la proliferación, diferenciación y actividad funcional de los linfocitos B (producción de anticuerpos).
¿De dónde vienen los linfocitos B en la zona cortical? Vienen aquí de la médula ósea roja con flujo sanguíneo. Los linfocitos B que ingresan al ganglio linfático están expuestos a antígenos fagocitados por macrófagos, antígenos retenidos en receptores de células dendríticas y linfocinas secretadas por T-helper. Bajo la influencia de todas estas influencias, los linfocitos B experimentan transformación blástica, proliferación y diferenciación dependiente de antígeno. Como resultado de esta diferenciación, los linfocitos B se convierten en células efectoras: células plasmáticas y células de memoria. Las células plasmáticas son capaces de producir anticuerpos dirigidos a destruir aquellos antígenos bajo cuya influencia experimentaron una diferenciación dependiente de antígenos. Luego, las células plasmáticas y las células de memoria ingresan al torrente sanguíneo general a través de las vénulas poscapilares y, después de una breve circulación, pasan al tejido conectivo. En el tejido conjuntivo, cada uno realiza su propia función: las células plasmáticas secretan anticuerpos y las células de memoria, al encontrarse con un antígeno familiar, se diferencian de las células efectoras y entran en una reacción inmunitaria destinada a destruir este antígeno.
Por lo tanto, los ganglios linfáticos son zona de linfocitos B.Si un antígeno ingresa al cuerpo, los nódulos se desarrollan en etapas.
Hay 4 etapas de desarrollo. En la etapa I, se forma un centro de luz; en este centro, los linfoblastos experimentan división mitótica.
En la etapa II, este centro se expande; en una sección de un ganglio linfático, hay hasta 10 divisiones mitóticas.
En el estadio III, aparece una corona alrededor de los centros claros de los nódulos, que consta de pequeños linfocitos; el número de células que se dividen disminuye, el centro de luz se estrecha.
En el estadio IV, las células en división son únicas; alrededor de un estrecho centro de luz hay una corona, que consiste principalmente en células de memoria.
El ciclo completo de cambios desde el momento en que ingresa el antígeno hasta el inicio de la etapa IV (etapa de reposo) dura de 2 a 3 días. Una semana después de que el antígeno ingresa al cuerpo, los cordones cerebrales se expanden, aumenta la cantidad de linfocitos y plasmocitos en los senos paranasales del ganglio linfático. Si los antígenos no ingresan al cuerpo (condiciones estériles), entonces no hay centros de luz en los ganglios linfáticos.
zona paracortical (paracorteza) se encuentra entre los ganglios linfáticos y los cordones cerebrales. Esta zona contiene células interdigitadas, linfocitos T y linfoblastos T. Las células interdigitadas se denominan así porque tienen procesos que penetran entre los extremos de los procesos de las células vecinas interdigitadas. Las células interdigitalizadas son macrófagos que han perdido la capacidad de fagocitosis, contienen orgánulos poco desarrollados de importancia general y tienen un citoplasma débilmente teñido. Las células interdigitadas producen glicoproteínas que estimulan la diferenciación de los linfocitos T, y las glicoproteínas ubicadas debajo de la membrana plasmática actúan como receptores que retienen antígenos que están involucrados en la diferenciación de los linfocitos T. En esta zona, hay una interacción cooperativa de células inmunocompetentes. Cuando se extirpa el timo (timectomía), la zona paracortical (dependiente del timo) se expresa mal.
Así, la zona paracortical es zona de linfocitos T, o zona dependiente del timo.
La médula de los ganglios linfáticos más ligero, formado como resultado del entretejido de los cordones cerebrales. El estroma de la médula también es tejido reticular. La composición de los cordones cerebrales incluye células plasmáticas. Los linfocitos B, los macrófagos y las células reticulares, es decir, los cordones cerebrales son la zona de los linfocitos B. Los capilares sanguíneos corren en los cordones del cerebro.
Senos de los ganglios linfáticos. Entre la cápsula y los ganglios linfáticos hay un seno subcapsular (sinus subcapsularis), entre las trabéculas y los ganglios linfáticos hay senos perinodulares (sinus perinodularis) que se extienden desde el seno subcapsular, entre las trabéculas y los cordones cerebrales hay senos cerebrales (sinus medullaris) que se extiende desde los senos perinodulares.
Los senos están revestidos de células reticuloendoteliales, es decir, células endoteliales especiales similares a los reticulocitos. Entre las células reticuloendoteliales hay células costeras - macrófagos. Estos macrófagos fagocitan los antígenos que se encuentran en la linfa que fluye en los senos paranasales. Las células reticuloendoteliales de los senos en la superficie de la pared adyacente a la cápsula y las trabéculas se encuentran en la membrana basal, y en la superficie que mira hacia los ganglios linfáticos y los cordones cerebrales, se encuentran en la malla de fibras reticulares que trenzan los ganglios linfáticos y cordones cerebrales. Esto contribuye al flujo de linfocitos y células plasmáticas desde los nódulos y los cordones cerebrales hacia la luz de los senos paranasales.
Funciones de los ganglios linfáticos .
La función hematopoyética, que consiste en la diferenciación de linfocitos dependiente de antígenos, lo que da como resultado la formación de células efectoras y células de memoria implicadas en reacciones inmunitarias, es una función de defensa inmunitaria.
Función protectora, que consiste en que los macrófagos fagocitan bacterias, fragmentos de células, antígenos que se encuentran en la linfa que circula por los senos paranasales.
Además, la linfa está enriquecida con linfocitos. La linfa se deposita en los ganglios linfáticos.
Los ganglios linfáticos del mesenterio del intestino participan en el metabolismo de los lípidos que se absorben capilares linfaticos vellosidades y transportado a los ganglios linfáticos.
Los ganglios linfáticos o ganglios linfáticos son órganos del sistema linfático humano. Son ovalados, redondos y, a veces, en forma de cinta y se distribuyen por todo el cuerpo humano, incluidas las axilas, y están interconectados por vasos linfáticos. Sus tamaños van desde 0,5 hasta 50 mm, pintadas en color rosado. Los ganglios linfáticos son el sitio de concentración de B, T y otras células inmunitarias.
Los ganglios linfáticos juegan el papel de trampas y filtros para partículas extrañas, son necesarios para funcionamiento normal sistema inmunológico humano. En los ganglios linfáticos hay una densa concentración de glóbulos blancos (linfocitos y macrófagos).
Los ganglios linfáticos son de importancia clínica. Se inflaman o agrandan varias enfermedades tanto infecciosas como oncológicas. La importancia del estado de los ganglios linfáticos para determinar la presencia de cáncer es importante.
Los ganglios linfáticos no son parte del sistema humano para combatir enfermedades asociadas con el envenenamiento. sustancias toxicas, estas tareas son realizadas por los riñones y el hígado.
Los ganglios linfáticos inflamados se diagnostican con una biopsia. Las enfermedades que causan inflamación de los ganglios linfáticos tienen un cierto curso y localización.
La estructura de los ganglios linfáticos.
Los ganglios linfáticos están rodeados por cápsulas fibrosas y dentro de la cápsula fibrosa se extiende para formar una trabécula. La sustancia del ganglio linfático se divide en sustancia cortical y médula, rodeada por todas partes por la corteza, con la excepción del hilio, donde la médula entra en contacto con la superficie.
Las fibras reticulares delgadas, la elastina y las fibras reticulares forman una estructura de malla de soporte: una red reticular dentro del nódulo, en la que los glóbulos blancos están densamente concentrados como folículos en la corteza. En otros lugares, solo están presentes leucocitos aislados. La función de la red reticular no es solo dar soporte, su superficie sirve para la adhesión de células dendríticas, macrófagos y linfocitos. Promueve el metabolismo con la sangre a través de las vénulas endoteliales y también proporciona los procesos necesarios para la aparición y maduración de las células inmunitarias.
El ganglio linfático tiene un canal: el seno linfático, revestido con células endoteliales y reticulares, fibroblastos, que proporciona un flujo suave de linfa. Así, el seno subcapsular es un seno situado directamente en la cápsula y su endotelio es continuado por un vaso linfático aferente. Los senos linfáticos, junto con el parénquima, constituyen una única unidad morfológica y fisiológica. En el lado convexo, varios vasos aferentes ingresan a la cápsula, trayendo linfa hacia el seno ubicado debajo de la cápsula. Este seno se llama marginal o circular. Este seno es una formación extensa en forma de hendidura que cubre todo el parénquima del nódulo, que se encuentra debajo de la cápsula y se interrumpe solo en los puntos de penetración de las trabéculas en el parénquima. Justo allí, el seno marginal pasa a los senos intersticiales (intermediarios), que luego son los senos cerebrales, pasando a la parte cerebral del nódulo, donde ya no se controla su conexión posterior. A diferencia de los anteriores, los senos cerebrales son grandes y su estructura parece ser esponjosa debido a que sus células endoteliales son más altas. Los senos intermedios y marginales están en estrecho contacto con los folículos linfáticos, creando conexiones funcionales. Los senos cerebrales se fusionan en las puertas de los ganglios linfáticos, formando vasos eferentes que transportan la linfa.
Las paredes de los senos linfáticos están compuestas por células endoteliales, que se denominan litoriales. Algunas de estas células tienen forma de estrella y están equipadas con procesos que cruzan el seno transversalmente y conectan sus paredes opuestas como puentes. La acumulación de estas células es un filtro para la linfa que fluye a través de ellas.
La diferencia entre los senos linfáticos y los vasos sanguíneos es que tienen una membrana basal en el lado de las trabéculas y la cápsula, mientras que no existe tal membrana en el lado del parénquima del vaso linfático. Esto se puede ver cuando se observa con un microscopio electrónico.
Donde hay tejido linfático, las células endoteliales están fuertemente presionadas contra las fibras reticulares que reemplazan la membrana basal. El citoplasma de las células endoteliales de los senos consiste en un retículo endoplásmico vesicular delgado y liso, y las células ubicadas cerca del parénquima linfático consisten en lisosomas y retículo endoplásmico granular.
Ubicación de los ganglios linfáticos
Los ganglios linfáticos se encuentran a lo largo de los vasos linfáticos en grupos de hasta 10 ganglios, así como cerca de los vasos sanguíneos y las venas grandes.
Los ganglios linfáticos se ubican en grupos para crear una barrera contra la propagación de infecciones y células cancerosas. Se ubican, por ejemplo, en los siguientes lugares: pliegues de rodilla, región inguinal, pliegues cubitales, axilas. Los ganglios linfáticos, que se encuentran en el cuello, protegen los órganos de la cabeza y los órganos circuncervicales de infecciones y tumores.
Ganglios linfáticos agrandados
Un aumento en los ganglios linfáticos indica la presencia de una enfermedad en el área donde se encuentra el ganglio. La mayoría de las veces esto se asocia con enfermedades infecciosas, y rara vez con tumores.
Linfadenitis aguda: la inflamación de los ganglios linfáticos ocurre cuando hay proceso purulento. Los principales síntomas son un aumento en el ganglio linfático, dolor cuando se siente. Durante el proceso inflamatorio sobre el ganglio linfático inflamado, la piel puede enrojecerse. Es posible que se requiera una apertura quirúrgica, si no se realiza a tiempo, el ganglio linfático puede romperse y su contenido penetra en los tejidos circundantes: se desarrolla flemón.
En los niños con tuberculosis se produce un aumento de los ganglios linfáticos, que es uno de los síntomas de la enfermedad.
Además, un aumento en los ganglios linfáticos puede estar asociado con una enfermedad como " rasguño de gato". En el que un microbio penetra a través de un rasguño: Bartonella, que es portada por los gatos.
Asimismo, un aumento de los ganglios linfáticos está asociado a enfermedades respiratorias y es consecuencia de una respuesta excesiva del sistema inmunitario a la infección.
Con la sífilis, se produce un aumento de los ganglios linfáticos en la ingle después de la aparición de una úlcera en los genitales.
Un aumento prolongado en varios ganglios linfáticos puede indicar la presencia de VIH, brucelosis, mononucleosis, listeriosis.
En los tumores, un aumento de los ganglios linfáticos puede estar asociado con la diseminación de metástasis, así como con el desarrollo de un tumor directamente en el ganglio linfático (linfogranulomatosis, linfosarcoma).
El parénquima de los ganglios linfáticos está impregnado de una densa red de canales (hendiduras): senos linfáticos, a través de los cuales la linfa que ingresa al ganglio fluye desde el seno subcapsular (marginal) hasta el portal. Directamente debajo de la cápsula del ganglio, entre este y el parénquima, hay un seno subcapsular (marginal), en el que fluyen los vasos linfáticos aferentes. Desde el seno subcapsular hasta el parénquima del ganglio, a lo largo de las trabéculas capsulares, salen los senos intermedios de la cortical y la médula. Estos últimos alcanzan la puerta del ganglio linfático (engrosamiento hiliar) y desembocan en el seno porta, desde donde se originan los vasos linfáticos eferentes. El seno subcapsular (marginal) también fluye hacia el seno portal, cubriendo el parénquima del órgano a lo largo de la periferia (debajo de la cápsula) y terminando en la región de la puerta del nódulo. Los senos intermedios de la sustancia cortical penetran profundamente en el órgano entre los nódulos linfoides, a veces se doblan y cubren el nódulo de un lado u otro (el seno cortical nodular redondo). Situados a lo largo de las trabéculas capsulares, los senos corticales, por un lado, son adyacentes al tejido conjuntivo de las trabéculas y, por otro lado, al parénquima linfoide de la sustancia cortical.
En la médula, los senos paranasales se ubican de dos maneras.
Algunos senos cerebrales intermedios se encuentran entre cordones pulposos adyacentes (seno interpulmonar). Otros están (en un corte histológico) entre los cordones pulposos de un lado y la trabécula hiliar del otro (seno trabecular). Los senos de la médula son más anchos que los senos subcapsulares y corticales. Las paredes delgadas de los senos desde el lado de su luz están revestidas con células endoteliales (costeras) engrosadas. El seno marginal del lado de la cápsula, así como las paredes de los senos corticales, cerebrales y portales, adyacentes a las trabéculas y al engrosamiento portal, tienen una capa continua de células costeras. Las paredes de los senos paranasales, orientadas hacia el parénquima linfoide, están construidas de tal manera que a través de ellas desde la cortical y la médula hacia la linfa y hacia direccion contraria los linfocitos, los macrófagos y otras células que se mueven activamente pueden penetrar fácilmente. En el lumen de los senos paranasales hay una red finamente enrollada formada por células y fibras reticulares. En los bucles de esta red, pueden permanecer partículas extrañas, cuerpos microbianos y células tumorales que ingresan al ganglio linfático junto con la linfa.
Usando microscopía de barrido de preparaciones corrosivas y nativas de V.
K. Shishlo y A. A. Mironov (1990) demostraron que la red de células reticulares y sus procesos, así como las fibras que existen en los senos paranasales, sirven para crear turbulencias en el flujo linfático. Esto facilita el filtrado de la linfa a través de la red dentro de los senos paranasales. Con respecto a las células que recubren las paredes de los senos paranasales, hay dos opiniones. Varios autores consideran estas células como células próximas al endotelio, siendo, por así decirlo, una continuación del endotelio de los vasos linfáticos aferentes. Según otro punto de vista, estas células son células reticulares modificadas con capacidad de fagocitosis. N. Siegmund (1923) dio a estas celdas el nombre de "costeras". Y. Mori y K. Lennert (1969) llamaron a las células sinusales retoteliales. En la estructura fina de las células retoteliales, se notan diferencias según su localización: cerca del seno marginal, senos intermedios de la corteza y la médula. En particular, L. V. Chernyshenko (1966) indica que el endotelio del seno marginal de los ganglios linfáticos parietales de la cavidad abdominal en fetos y recién nacidos tiene una estructura celular, mientras que en los ganglios pancreáticos superiores y anteriores y gástricos superiores tiene un "sincitio". " estructura.
La mayoría de las células costeras (retoteliales) contienen filamentos delgados a lo largo del eje longitudinal de las células. Las células vecinas están conectadas por desmosomas. A diferencia de las células del seno marginal, las células costeras de los senos intermedios contienen una gran cantidad de orgánulos. Contienen numerosas vesículas del retículo citoplasmático liso y túbulos largos del retículo citoplasmático granular.
Como lo muestran Yu.I.Borodin y V.N.Grigoriev (1983), las células que forman la pared exterior del seno están interconectadas adhiriéndose entre sí y formando contactos complejos, como los interdigitales. Al mismo tiempo, se producen manchas y zonas de obliteración a lo largo de los contactos, lo que indica la densidad de estos contactos. La forma de los núcleos celulares de la pared exterior del seno marginal es ovalada o alargada, y en ocasiones incluso en forma de bastón. La cromatina es de grano fino y se distribuye principalmente a lo largo de la periferia del núcleo. La membrana citoplasmática forma pliegues frente a la luz del seno y al tejido conjuntivo adyacente. A veces, los linfocitos "encajan" muy cerca de las células del revestimiento exterior del seno, y sus prolongaciones ingresan a las invaginaciones de las células de la pared del seno. Los orgánulos se distribuyen uniformemente por todo el citoplasma. Las mitocondrias suelen tener forma redonda u ovalada, con un pequeño número de crestas. Los retículos citoplasmáticos granulares y no granulares están poco desarrollados, predomina el retículo granular. Hay un gran número de ribosomas y políticas libres en forma de cadenas o rosetas. Los lisosomas son pocos, densos en electrones, redondeados, con contenido granular. En la zona perinuclear del citoplasma existe un complejo lamelar formado por cisternas aplanadas y vesículas. Las vesículas se encuentran en todo el citoplasma de la célula, a veces se fusionan, formando vacuolas. Por lo tanto, las células que forman la pared externa del seno se asemejan al endotelio en ultraestructura.
al estudiar pared interior sinus V. N. Grigoriev identificó 3 tipos de células que difieren entre sí en características morfológicas. Las células del primer tipo tienen procesos y se ponen en contacto entre sí por medio de una simple unión de procesos. En la luz entre los procesos citoplasmáticos hay haces de fibras reticulares. Los núcleos (raramente incluidos en el corte) ocupan la parte central de la célula. Su forma es ovalada, redondeada, a veces irregular; la cromatina es de grano fino; Los orgánulos se encuentran principalmente en la zona del pericarion. Las mitocondrias son redondas u ovaladas, con pocas crestas. El retículo citoplasmático granular está bien desarrollado. El citoplasma de estas células, así como el de las células que forman la pared externa del seno, contiene una gran cantidad de ribosomas y polisomas libres. Algunos lisosomas parecen formaciones redondeadas densas en electrones con contenido granular. El complejo lamelar se encuentra más a menudo cerca del núcleo. La actividad pinocitótica en estas células no se expresa. La pared interna del seno marginal, representada por células tipo 1, tiene poros en los que a veces son visibles los linfocitos. Las células del segundo tipo, que recubren la pared interna del seno, en contraste con las células del primer tipo, son aplanadas y alargadas. Se contactan entre sí por un simple ajuste de los procesos. Los núcleos son ovalados o en forma de varilla, la cromatina es de grano fino. Los orgánulos de estas células, en contraste con los orgánulos de las células tipo 1, están poco desarrollados: son visibles mitocondrias únicas, un retículo citoplasmático liso y ribosomas libres. Numerosas vesículas de pinocito. Rara vez se encuentran células del tercer tipo; están ubicados directamente en la luz de los senos paranasales cerca de las células del segundo tipo y están en estrecho contacto con ellas. Tienen una forma ovalada, los procesos de estas células forman estrechos contactos con el cuerpo de su propia célula. Los contornos de la membrana nuclear son enrevesados, lo que indica un aumento en la superficie del núcleo. La cromatina es de grano fino. Los orgánulos se desarrollan de la misma manera que en las células tipo 1. Sin embargo, a diferencia de estas últimas, las células tipo 3 contienen muchos lisosomas y vesículas pinocíticas.
El tamaño de las células costeras de los senos cerebrales supera significativamente el tamaño de las células costeras de la pared interna del seno marginal. Por lo tanto, las proporciones de volumen de estos últimos son notablemente mayores que las de las células costeras de los senos cerebrales. Con una disminución en la función del sistema inmunológico en animales (e incluso con inmunodeficiencia), como G. Sainte-Marie y F.-S. Peng (1990), los senos subcapsulares se expanden. Parece que tienen pocos linfocitos. Al mismo tiempo, los senos de la médula están llenos de numerosos macrófagos. Los senos se caracterizan por la penetración de células en la zona nodular, y esto ocurre en el contexto de la atrofia de los nódulos linfoides, la presencia de los llamados linfocitos oscuros, que se consideran células dañadas. Según N. Chizini-Garcia, S. R. Machardo (1992), aquí en los senos paranasales también hay mastocitos que se distinguen por el tamaño heterogéneo, la forma y las propiedades tintóreas de los gránulos. Es posible que contengan heparina, pero es menor en los senos cerebrales que en el resto del ganglio linfático.
En el funcionamiento de los ganglios linfáticos, que pasan la linfa a través de sí mismos y actúan como filtros biológicos, intervienen todas sus estructuras: corteza y médula, numerosos senos linfáticos ramificados en el parénquima. En este sentido, son de interés los datos sobre los procesos dinámicos en los ganglios linfáticos y el papel de los linfocitos T y B en ellos. Según P. V. Pigarevsky (1991), normal componentes estructurales Las zonas T y B de los ganglios linfáticos viscerales y somáticos están compuestas en un 90-95 % por linfocitos de tamaño pequeño y mediano y células reticulares, y solo el 5 % son formas celulares de la serie plasmacítica. En la zona T, el número de T-helpers es mayor que el de los T-supresores, pero esta proporción puede cambiar durante el día. Normalmente, dentro de las zonas T y B de los ganglios linfáticos somáticos y viscerales, no se produce ninguna reestructuración debido a la formación de un clon de células sensibles al antígeno necesarias para el desarrollo de una respuesta inmunitaria. En la zona T, rara vez se observaron células en división mitótica (del 0,02 al 0,06 %, resultó que cambios relacionados con la edad este indicador no se ve afectado). A. Yu. Letyagin (1991) cree que la estructura espacio-temporal diaria del ganglio linfático consiste en complejos morfofuncionales rítmicamente temporales. El complejo "A" seleccionado se caracteriza por un aumento en el número de estructuras linfoides del ganglio linfático como resultado de la recirculación y proliferación de linfocitos. El complejo "B" se caracteriza por una disminución significativa y rápida en el número de linfocitos en el órgano debido a la emigración de estas células a la circulación periférica en el contexto de la activación de factores dinámicos del microambiente y la actividad de los sistemas de drenaje que aseguran el descarga de líquido del órgano. El complejo "B 2" se caracteriza por un equilibrio inestable de células y fluidos en el órgano. Al mismo tiempo, hay un aumento en la migración de linfocitos bajo la influencia activa de factores del microambiente. Todos estos datos se presentan de forma más completa en la monografía de Yu.I. Borodin, V.A. Trufakin et al. (1992), postulando que “el proceso de migración y recirculación de elementos linfoides, incluso en ausencia de inmunización, no es aleatorio. Los linfocitos "conocen" los límites de su área de acción, su ruta de movimiento en el sistema linfoide y los tejidos periféricos. Tal orden surge en el proceso de ontogénesis tanto de un solo linfocito como de todo el sistema linfoide, y se correlaciona con la capacidad de sintetizar y transportar receptores específicos”.
I. T. Gegin y A. I. Krayushkin (1991) toman los nódulos linfoides con territorios adyacentes, cordones pulposos como una continuación de los nódulos en lo profundo de la médula y trayendo vasos linfáticos como una unidad estructural y funcional en los ganglios linfáticos. En la monografía de Yu. I. Borodin, M. R. Sapin et al. (1992) resumieron los intentos individuales de subdividir de alguna manera el tejido del ganglio linfático en unidades estructurales. Así, G. Sainte-Marie et al. (1984, 1990), Yu. E. Vyrenkov, V. K. Shishlo y Yu. G. Antropova (1993) no se destacan de manera muy convincente como unidad estructural compartimento de los ganglios linfáticos. Este sector, que según los autores incluye la parte central del ganglio linfático, correspondiente a la región paracortical, en este sector se encuentran varios nódulos linfoides y cordones pulposos. parte central llamado unidad o dominio T. Este último está formado por elementos del estroma, varias poblaciones de linfocitos T y un pequeño número de linfocitos B que migran a sus respectivos dominios B. El dominio T se extiende hacia el espacio extranodular, donde ocurre la migración selectiva de linfocitos a través de las paredes de las vénulas con endotelio alto. Yu. E. Vyrenkov, V. K. Shishlo y Yu. G. Antropova (1993) indican que cada compartimento está "dedicado" a un solo vaso linfático aferente, 1-6 nódulos linfoides.
A. I. Krayushkin (1994) escribe sobre la presencia en los ganglios linfáticos de partes estructurales y funcionales correspondientes a órganos y regiones del cuerpo que son heterogéneas en función y estructura. Al examinar los ganglios linfáticos de conejos de diferentes edades, mostró que en el ganglio linfático mesentérico central (visceral), que recibe linfa tanto del intestino delgado como del grueso, los tintes introducidos en las paredes de estos órganos, al ingresar al ganglio linfático, llenar los senos paranasales, cada uno en su parte. Este autor también señaló que los vasos linfáticos de intestino delgado acérquese a un lado (izquierda) del ganglio linfático mesentérico, y desde el colon, al lado opuesto (derecho) de este ganglio. Yu. I. Borodin (1994) también escribió sobre la correspondencia de los vasos linfáticos aferentes con una cierta parte del parénquima del ganglio linfático. Según A. I. Krayushkin, los senos linfáticos en los que ingresa la linfa desde las paredes del intestino delgado son más anchos que los senos que "pertenecen" al intestino grueso. Este hecho también indica una mayor carga de transporte de los senos de la parte del "intestino delgado" del ganglio linfático en comparación con la parte del "colon". Esta suposición es consistente con los datos de N. Reed (1989), quien encontró que el intestino delgado tiene una mayor capacidad de absorción, y en sus paredes, gran cantidad sustancias que en el intestino grueso.
La misma presencia de partes morfofuncionales se observa en los ganglios linfáticos poplíteos somáticos, a los que fluye la linfa desde los tejidos superficiales y profundos de la extremidad. Como lo muestra A. I. Krayushkin (1994), la linfa de la piel y tejido subcutáneo, y en el otro, a través de vasos profundos, desde las cápsulas de las articulaciones, músculos, tendones, fascia de la extremidad pélvica del conejo. El tinte inyectado en los músculos y otras estructuras de la extremidad ubicadas profundamente se envía a través de los vasos linfáticos profundos al ganglio linfático poplíteo y se encuentra en los senos paranasales de la parte anterior de este ganglio. Los senos linfáticos en la parte posterior del ganglio linfático están llenos de colorante que proviene de los tejidos superficiales (piel y tejido subcutáneo). El autor señala que los senos intermedios del bulbo raquídeo son más anchos en la parte del ganglio linfático a la que fluye la linfa desde los tejidos superficiales de la extremidad. Se puede suponer que el parénquima linfoide adyacente a los senos linfáticos, que reciben linfa de las paredes del intestino delgado (ganglios mesentéricos) o de la piel y tejido subcutáneo (ganglios poplíteos), experimentará una mayor carga funcional que el parénquima de la otra parte de estos nodos.
Los datos obtenidos por AI Krayushkin confirman esta suposición. Al investigar el tejido linfoide de los ganglios linfáticos, en el que fluyen los vasos linfáticos, que transportan linfa de órganos funcionalmente diferentes, A. I. Krayushkin descubrió hechos interesantes. En esa parte del ganglio linfático donde la linfa fluye desde las paredes del intestino delgado, el parénquima linfoide tiene mayor indicadores cuantitativos que en la otra parte del nódulo, interconectado con el intestino grueso. En la parte del "intestino delgado" del ganglio linfático mesentérico, los nódulos linfoides (zona B) son más grandes, su diámetro varía de 275 a 750 micrones (promedio de 467 micrones). El área ocupada por los nódulos linfoides es el 20,3% del área de corte del ganglio mesentérico. Una gran área en las secciones del nudo también está ocupada por hebras pulposas. En estas partes del ganglio linfático mesentérico, también hay un mayor contenido de formas blásticas de células, linfocitos grandes y plasmocitos, lo que indica un aumento en las funciones linfocito e inmunopoyéticas aquí. En la parte "colónica" del ganglio linfático, el diámetro de los nódulos linfoides es mucho más pequeño: 349 μm (200-625 μm), y el área de los nódulos linfoides en la sección del ganglio es solo del 13,1%. En esa parte del nódulo mesentérico, donde entra la linfa del intestino delgado, la zona paracortical (zona T) ocupa 3 veces menos área (8,1%) que en la parte a la que fluye la linfa del intestino grueso (25,9%). ) .
En el ganglio poplíteo somático, también hay diferencias en la organización del parénquima linfoide en las partes hacia las que fluye la linfa. diferentes regiones extremidades En esa parte del ganglio, a la que se acercan los vasos linfáticos de la piel y el tejido subcutáneo, hay parámetros morfométricos más altos de nódulos linfoides y cordones pulposos (zonas B). El diámetro de los nódulos linfoides en esta parte era de 375 μm (193-646 μm) y su área era del 15,2% de la sección total del nódulo. En otra parte del ganglio linfático poplíteo, al que fluye la linfa desde los tejidos profundos de la extremidad (músculos, tendones, cápsulas articulares), el diámetro de los nódulos linfoides era inferior a 302 micras (123-492 micras), y su área en el corte ocupaba la mitad del área -7, 9%. La zona T paracortical resultó ser más ancha en la parte donde la linfa ingresa desde los tejidos profundos de la extremidad.
A. I. Krayushkin explica tales diferencias en dos zonas morfofuncionales adyacentes del ganglio linfático poplíteo por un efecto más fuerte y más constante en la piel de "antígenos ambiente externo". No es casualidad que en la piel se encuentren nódulos linfoides perivasculares, que son estructuras inmunitarias de la piel. El desarrollo predominante de nódulos linfoides y cordones pulposos (zonas B) en la parte del ganglio linfático mesentérico central, donde la linfa ingresa desde el intestino delgado, y en el ganglio poplíteo, al que se envía la linfa desde la piel y el tejido subcutáneo. indica la formación de respuestas inmunes en estas partes de tipo humoral. Estas reacciones producen anticuerpos. Muchos investigadores han demostrado que los nódulos linfoides contienen complejos inmunes. Según V. I. Novikov y A. A. Vlasov (1989), S. T. Lee, F. Paraskevas y J. Maeba (1985), S. T. Lee y F. Paraskevas (1988), los complejos específicos de antígeno incluyen determinantes de antígeno, moléculas de histocompatibilidad de clase II e inmunoglobulina. determinantes Los macrófagos y los linfocitos T son necesarios para la formación de dicho complejo.
V. I. Novikov et al. (1990) notan que las células de los ganglios linfáticos periféricos después del contacto con el antígeno in vitro secretan factores específicos de antígeno. Estos factores inhiben la migración de macrófagos y aumentan su actividad funcional, y también contribuyen a la diferenciación final de las células progenitoras de la serie granulocitos-macrófagos a lo largo de la vía monocítica. Según estudios de S. K. Drinker, N. Field y N. Ward (1934), es en los ganglios linfáticos donde se puede encontrar hasta el 99% del material antigénico. Por lo tanto, las células activadas por antígeno se acumulan selectivamente en estos nodos que, como lo muestran V. I. Novikov y A. A. Vlasov (1989), V. I. Novikov, A. A. Vlasov e I. G. Sidorovich (1991), participan en reacciones inmunológicas contra este antígeno. En el punto máximo de la respuesta inmunitaria, las células de los ganglios linfáticos secretan un antígeno, un factor humoral específico que potencia la génesis de anticuerpos en el período productivo al suprimir la función de los supresores de células T. Los autores concluyen que las células de los ganglios linfáticos "participan no solo en la acumulación y el procesamiento del material antigénico capturado en una forma que, con el flujo de la linfa y la sangre, alcanza los órganos centrales de la inmunidad (en particular, médula ósea), provocando cambios observados en la producción de inmunomoduladores en la misma, pero también en la regulación de etapas posteriores en el desarrollo de la respuesta inmune, incluido el periodo productivo de esta reacción. N. van Rooijen (1987) sugiere que existe una forma de migración de células que se diferencian en los ganglios linfáticos en células formadoras de anticuerpos. En una respuesta inmune independiente del timo, las células B reactivas al antígeno reciben la información necesaria de los macrófagos presentadores de antígenos en la zona internodular de la parte periférica de la corteza. M. Dohrzanski y T. Yang (1991) mostraron que los linfocitos T de varios ganglios linfáticos difieren en número y características funcionales.
GV Kovalevsky (1992) identificó 3 etapas de cambios en la morfología funcional de la respuesta inmune del ganglio linfático a los antígenos dependientes del timo. 1. Etapa redistributiva - 1er día de la respuesta inmune. Los ganglios regionales estimulados por antígenos producen un agente humoral que promueve la liberación de linfocitos T de los ganglios linfáticos no regionales. Al mismo tiempo, aparecen focos de devastación de la zona paracortical en este último en el contexto de la linfocitosis más aguda de los senos paranasales. En la médula ocurre la autofagia de las células plasmáticas y se forman macrófagos esféricos gigantes. 2. Etapa proliferativa - el 3er día de la respuesta inmune, el momento de máxima eliminación de antígenos. Algunas áreas de la zona T pierden su composición habitual de células pequeñas y se convierten en campos de "grandes células pironinofílicas". Se supone que todas las subpoblaciones de linfocitos T específicos (auxiliares, amplificadores, supresores, asesinos, etc.) surgen de ellos. Por lo tanto, en esta etapa se desarrolla la autorregulación de la respuesta inmune en la zona paracortical. 3. La etapa final, relativa a la morfogénesis, se manifiesta en el día 5-6 de la respuesta inmune. Hay una movilización masiva de linfocitos hacia los senos de los ganglios linfáticos regionales. La recirculación involucra linfocitos T no sensibilizados, que además reciben información antigénica a través de la interacción de contacto (peripolesis) con macrófagos sinusales. Como señala G. V. Kovalevsky, los linfocitos T sensibilizados son extraídos del endotelio de las vénulas poscapilares de los ganglios linfáticos no regionales y pueblan allí los lugares que surgieron en la primera etapa. Las "ondas" de los centros de reproducción se observan cuanto más tiempo, mayor fue la dosis de antígeno.
Según G. G. Csanaky, V. Kalasz y T. Rahr (1991), después de la estimulación antigénica, tanto el número de venas con endotelio alto como la masa del ganglio linfático aumentan rápidamente.
G. Sainte-Marie y F. S. Peng (1985), T. S. Smirnova (1992) creen que las células ubicadas en la parte superficial de su corteza (directamente debajo del seno marginal) contribuyen a la transferencia de antígenos de la linfa entrante al parénquima de la ganglio linfático, llamado zona marginal. Según G. Sainte-Marie y F. S. Peng (1985), el estroma de la zona subcapsular está representado por células reticulares fibroblásticas que forman una red en forma de bucle. J. E. Veldman, F. J. Heuning e I. Molendar (1978), M. Dobashi, K. Terashima y J. Imai (1982) indican que en la zona subcapsular, los linfocitos se transforman en inmunoblastos, plasmablastos y células plasmáticas, independientemente de la presencia de células T. regulación y formación de centros de reproducción.
Según T. S. Smirnova (1992), este proceso puede continuar independientemente del estado de los centros de reproducción, donde el desarrollo y transformaciones celulares ocurren mucho más rápido que en otras estructuras del ganglio linfático. T. S. Smirnova también estudió los cambios rítmico-temporales en la población de células linfoides en la zona marginal y comparó los datos obtenidos con los patrones de la composición celular en las zonas dependientes de T y B del mismo nodo. El análisis de los resultados mostró la ausencia de "componentes comunes", que sirvieron de base para el enunciado sobre la "autonomía funcional relativa" de la zona marginal. El autor cree que esta zona es el sitio de entrada al parénquima del ganglio linfático de pequeños linfocitos y mastocitos de la linfa que ingresa al ganglio.
En muchos estudios se indica un cierto ritmo en la estructura de las formaciones linfoides de los ganglios linfáticos. En la literatura científica, hay evidencia de que la masa de ganglios linfáticos somáticos y viscerales cambia durante el día. Según Yu.P.Hussar (1969, 1975), la masa máxima de los ganglios linfáticos de las ratas blancas cae en 18 horas y la mínima en 3 horas. densidad máxima la ubicación de los linfocitos en los ganglios linfáticos axilares, inguinales y mesentéricos también cae en horas 18. Yu.I.Borodin et al. (1992) revelaron biorritmos de 24 horas de composición celular en la meseta cortical de los ganglios linfáticos inguinales de ratones CBA. En la meseta cortical de los ganglios linfáticos mesentéricos, se observa un ritmo de cambios de 12 horas en el número de células linfoides: un aumento por la noche y el día, una disminución por la mañana y por la noche. Los autores explican estos hechos por la actividad motora durante las comidas por la noche y tiempo de día. La disminución del número de células en la meseta cortical se produce durante el sueño de los animales. En esta meseta de ganglios linfáticos de bifurcación, el contenido máximo de células se observa durante la noche y el mínimo durante el día.
Yu. I. Borodin et al. considerar que el aumento del número de células en la fase nocturna está asociado a la migración de células en este momento del día desde el timo, lo que concuerda con las conclusiones de Yu. P. Khussar (1975). Esto no excluye la posibilidad de redistribución de células en los propios ganglios linfáticos, así como su liberación al torrente sanguíneo en condiciones de actividad motora animal. También se trata de la recirculación de los linfocitos existentes en el organismo. Es imposible no notar, como creen los autores, la inestabilidad de los ritmos circadianos en la composición celular de la meseta cortical, que depende de una serie de razones: la posición de los ganglios linfáticos en el cuerpo, los niveles de proliferación, celular recirculación, etc
Se encontró un patrón diferente de ritmos celulares en la zona paracortical dependiente del timo de ratones CBA. En los ganglios linfáticos inguinales, el número máximo de células de la serie linfoide cae durante el día y la noche. En la zona paracortical de los ganglios linfáticos mesentéricos, se observa una disminución en el número de células durante el período de actividad motora de los animales durante la noche. Se observó un aumento en el número de células durante el día también debido a la actividad motora. Los autores consideran la dinámica diaria de la composición celular en relación con las diferentes direcciones de migración celular que involucran el timo, el bazo, otros grupos de ganglios linfáticos y el torrente sanguíneo. El sincronismo de los biorritmos diarios de la composición celular en la meseta cortical y la zona paracortical enfatiza la relación funcional de varias estructuras linfoides en estos órganos. Según N. Metcalf, S. Youngberger y W. Metcalf (1971), en ratas la tasa de recirculación de los linfocitos B es menor que la de los linfocitos T. Según los datos de A. Ya. Friedenshtein y E. A. Luria (1980), en la sustancia cortical, donde hay muchas vénulas y capilares sanguíneos Los linfocitos B migran desde la sangre a través de las paredes de estos microvasos hacia el parénquima de los ganglios linfáticos, hacia sus nódulos linfoides y cordones pulposos. Hay evidencia en la literatura de que hasta el 0,2% de todos los linfocitos B se encuentran en estas estructuras de los ganglios linfáticos.
La migración de linfocitos desde los nódulos linfoides también obedece a las leyes de los ritmos diarios. En los nódulos linfoides de los ganglios de bifurcación se encontró un ritmo de actividad celular de 12 horas con aumento del número de células en estas estructuras en horas de la mañana y de la tarde, entre las cuales hubo una ligera disminución de las células contenidas en a ellos. "Acumulación de células" en nódulos linfoides durante el día Yu. I. Borodin et al. (1992) explican la presencia de actividad mitótica en estas estructuras, así como la recirculación de linfocitos. Los autores creen que en las horas de la tarde, los linfocitos migran desde los nódulos linfoides de cualquier ganglio linfático, donde sea que se encuentren. Este proceso está asociado con el comienzo de la actividad motora vespertina de los animales. Se observó actividad motora tanto en las horas previas al amanecer como durante el día (alimentando a los animales). Los autores llegaron a la conclusión de que la formación de células en los nódulos linfoides de los ganglios linfáticos se observa durante el sueño de los animales y una disminución en su número durante el período de actividad física. En los animales que llevan un estilo de vida nocturno, el proceso de migración de linfocitos desde los nódulos linfoides se produce de forma natural durante la noche.
En las hebras pulposas de los ganglios linfáticos somáticos (inguinales) y viscerales (bifurcación y mesentéricos), se observa un ritmo de actividad celular de 12 horas. En horas de la mañana y de la tarde, el número de células de la serie linfoide en los cordones pulpares disminuye, y en el día y la noche aumenta, lo que no es consistente con los biorritmos de las células en los nódulos linfoides. Por lo tanto, Yu. I. Borodin et al. (1992) concluyen que los cordones pulpares no son estructuras completamente dependientes de B de los ganglios linfáticos. La dinámica de las células en los cordones pulposos es más similar a los mismos procesos en la meseta cortical y la zona paracortical, y los autores sugieren un papel acumulativo de los cordones pulposos en relación con las células de la serie linfoide. Un estudio biotecnológico de las características funcionales de los cordones pulpares reveló su relación con las zonas B y T de los ganglios linfáticos. La entrada de linfocitos en los cordones pulpares desde el exterior se observó en la primera mitad del día, y la inmigración - por la noche, cuando actividad física animales es muy alto.
En cuanto a los biorritmos en las estructuras linfoides de otros órganos del sistema inmunitario, Yu. P. Hussar, Yu. Teikes y X. Lyaene (1971), L. Scheving et al. (1972) indican cambios en la masa del bazo en ratas durante el día. Se observa un aumento en la masa durante las horas de descanso, la masa máxima se observa antes del amanecer y la mínima, durante el día y la noche.
Yu.P. Hussar (1975) observó aproximadamente la misma imagen de un aumento en el número de células de la serie linfoide en la mañana y una disminución en las horas de la tarde y el día en la pulpa blanca del bazo en ratones CBA machos. . Un aumento significativo en el número de linfocitos y un aumento en la densidad de su ubicación en la pulpa blanca del bazo se puede atribuir a la migración de estas células desde la médula ósea, donde en este momento el número de dichas células disminuye. Por la noche, también se observó un aumento de la actividad mitótica en el bazo.
Es posible que se produzca un aumento de la masa de tejido linfoide en el bazo en las primeras horas de la mañana y en la segunda mitad del día debido a la redistribución activa de las células linfoides en condiciones de cambios en las cargas funcionales de varios órganos durante el día. Se puede suponer que durante el día los linfocitos están más "ocupados" con el control de los antígenos alimentarios en las paredes de los órganos digestivos y en otros órganos que trabajan activamente durante el día. Como escriben Yu.I.Borodin et al. (1992), en las horas de la tarde y la noche, el "sistema experto" registra la liberación de células del bazo en la sangre y las zonas B-dependientes de los ganglios linfáticos. Es posible que el "control" de la sangre, que se realiza las 24 horas del día, alcance un máximo en las horas previas a la mañana. Como muestran G. Fernandes et al. (1976), durante el día en el bazo hay un número mínimo de linfocitos T con un contenido máximo simultáneo en la sangre periférica.
Los experimentos realizados por T. S. Smirnova al infligir lesiones en animales también indican cambios significativos en la composición celular de los ganglios linfáticos (en su zona marginal). Después de la lesión (en el día 1-2) en esta área, la cantidad de linfocitos pequeños aumentó en un 75%. Este aumento en l "/ 2 veces fue mayor que en otras áreas del ganglio linfático. T. S. Smirnova concluyó que los linfocitos llegan a la zona marginal desde el sitio de la lesión. El autor no explica de dónde provienen los linfocitos en el área de lesión y por qué es desde allí que ingresan al ganglio linfático. Como señala T. S. Smirnova, para el día 7-8 después de la lesión, la "densidad de población" de linfocitos en la zona marginal se normaliza. Al mismo tiempo, ella llama la atención sobre un comportamiento diferente de otras células de la serie linfoide, el número que en la zona marginal no aumenta, sino que, por el contrario, disminuye. Así, al tercer día después de la lesión, el número de linfocitos medianos disminuye en más de 3 veces, células en división mitótica - 6 veces, formas blásticas de células - 21/2 veces, células plasmáticas inmaduras - 3 veces, células plasmáticas maduras - 10 veces T. S. Smirnova también llama la atención sobre la presencia en el postraumático período en la zona marginal del ganglio linfático regional de los mastocitos, sobre el cual aumenta bruscamente al tercer día. El número de mastocitos también aumenta en el área de la lesión. Este sincronismo de la aparición de mastocitos en el sitio de la lesión y la zona marginal indica, en opinión del autor, un posible aumento de la intensidad del flujo sanguíneo local, estimulación de la actividad fagocítica de los macrófagos y presencia de quimiotaxis en los linfocitos. . Al mismo tiempo, T. S. Smirnova se refiere a los datos de N. A. Yurina y A. I. Radostina (1977) sobre el papel de los mastocitos en el cuerpo y J. Raud et al. (1989) sobre la ubicación periarterial de los mastocitos para ayudar a guiar la migración de leucocitos. En caso de trauma en la zona marginal, el ritmo de la población celular también cambia. Es cierto que la amplitud de las fluctuaciones y cambios identificados por T. S. Smirnova después de la lesión disminuye.
Sobre la base de los estudios realizados, T. S. Smirnova cree que en el plan funcional, la zona marginal ocupa una posición especial en el ganglio linfático. El autor cree que la zona marginal es el lugar de entrada de "linfocitos y mastocitos al parénquima de los ganglios linfáticos desde el canal linfático, tanto en condiciones normales como en respuesta al trauma quirúrgico". Por lo tanto, en la etapa actual de los estudios morfológicos, las observaciones cronobiológicas son muy prometedoras. Permiten comprender mejor los patrones de los procesos morfogenéticos a nivel celular, tisular y orgánico. Con respecto a los ganglios linfáticos, también resultó (en el experimento) que los biorritmos diarios del suministro de sangre al tejido linfoide son directamente consistentes con los ritmos diarios del suministro de sangre a las regiones del tejido. Los cambios locales en los ganglios linfáticos están influenciados por factores hormonales y nerviosos, la dinámica de la población diaria de mastocitos. Es posible que los mastocitos presentes en los tejidos perivasculares y linfoides, en los senos, cápsula y trabéculas de los ganglios linfáticos afecten tanto al microambiente como a la formación de la respuesta inmune. Los mastocitos no solo pueden formar grupos de 3-5 elementos, sino incluso estructuras "en forma de hongo" que comienzan en la región de la puerta del nódulo y continúan a lo largo de las trabéculas y los vasos hasta la meseta internodal cortical. Los detalles de la absorción de los gránulos de mastocitos liberados por las células reticulares aún no están claros. El autor cree que la zona marginal es el sitio de entrada al parénquima del ganglio linfático de linfocitos pequeños y mastocitos de la linfa que ingresa al ganglio.
La colocación de los primeros ganglios linfáticos se encontró en embriones humanos de 5 a 6 semanas de edad. Según V. A. Trufakin (1967), los primeros ganglios linfáticos mesentéricos comienzan a formarse en la octava semana de desarrollo intrauterino. V. A. Florensov (1983) descubrió que la colocación más temprana de los ganglios linfáticos ya se observa en embriones de 7 semanas. Los ganglios linfáticos formados anatómicamente se convierten en fetos de 9 semanas. M. A. Dolgova (1967), N. A. Zharikova (1979) y otros investigadores señalan esta edad (8-10 semanas), en la que comienza la formación de ganglios linfáticos. Cabe señalar que el momento de la formación de los ganglios linfáticos, según diferentes autores, por regla general, es diferente. Esto se puede entender considerando lo siguiente. En primer lugar, varios grupos de ganglios linfáticos sirvieron a menudo como objeto de estudio. En segundo lugar, no se tuvo en cuenta la salud de la madre que llevó estos embriones y fetos, las condiciones para obtener el material, etc.. En este sentido, uno debe estar de acuerdo con G. V. Kharlova (1963) en que la formación de ganglios linfáticos es una respuesta compensatoria a las condiciones de existencia del organismo.
Dado que los ganglios linfáticos están ubicados en las vías del flujo linfático de los órganos y tejidos, la aparición temprana de ganglios no es sorprendente. Ya en las primeras semanas del desarrollo del embrión, aparecen productos metabólicos en el líquido tisular, incluidas proteínas gruesas e incluso partículas de células destruidas formadas como resultado de la renovación celular. Dado que las células muertas, las proteínas gruesas y las posibles células mutantes son partículas extrañas, se requieren estructuras inmunitarias para su reconocimiento y destrucción. Tales estructuras son los ganglios linfáticos formados en el cuerpo del embrión, que son filtros biológicos en las vías de circulación de fluidos hacia el torrente sanguíneo. Es probable que la aparición en el cuerpo del embrión de productos metabólicos y partículas extrañas en la cantidad en que se vuelve peligrosa para el organismo en crecimiento sea el estímulo, el punto de partida para la formación de ganglios linfáticos. En diferentes regiones del cuerpo, se crea tal necesidad de aparición de ganglios linfáticos y sus grupos regionales en diferente tiempo. Por lo tanto, la aparición de pestañas de nodos en varios campos cuerpo humano se produce en varios períodos del desarrollo intrauterino, hasta el nacimiento e incluso después de él.
La formación de un ganglio linfático comienza con el aislamiento de un grupo de células mesenquimales cerca de un vaso linfático o entre vasos linfáticos adyacentes. Luego, en el proceso de desarrollo, el grupo de células se invagina en la luz del vaso linfático adyacente, cuya pared exterior da lugar a la cápsula del ganglio linfático resultante. La luz del vaso linfático se convierte aún más en el seno subcapsular (marginal). Los senos intermedios se desarrollan sobre la base de un plexo ramificado de vasos linfáticos, entre los cuales crecen hebras de tejido conjuntivo embrionario. En el futuro, aumenta el número de células mesenquimales en el rudimento del nodo. En el tercer o cuarto mes de desarrollo intrauterino, los linfocitos se determinan en los ganglios linfáticos en desarrollo.
A. Yasushi (1992) encontró que en los fetos, los nódulos linfoides comienzan a aparecer en la semana 16 de desarrollo debajo del seno subcapsular. A partir de la semana 18 se encuentran constantemente nódulos linfoides bien visibles. El autor explica la ausencia de centros de reproducción en los nódulos por la inmadurez de los tejidos y células dendríticas nodulares, así como por la falta de estimulación durante el desarrollo embrionario. Según J. Westerga y W. Timens (1989), se detectan células B de tamaño mediano en los nódulos linfoides y las secciones externas de la sustancia cortical de los ganglios linfáticos en los fetos. A partir de la semana 19, en algunos nodos se puede ver el borde delineado entre la futura cortical y la médula. Por lo tanto, los nódulos linfoides en los ganglios linfáticos ya aparecen en el período prenatal. P. Fusari (1965) observó la formación de nódulos en un feto de 51/2 meses, V. A. Florensov (1964), en la semana 34 del desarrollo intrauterino. Los centros de reproducción en los nódulos linfoides aparecen poco antes del nacimiento o poco después.
La diferenciación de los ganglios linfáticos continúa no solo en los recién nacidos, sino también en infancia. Los principales procesos de conformación relacionados con la edad en los ganglios linfáticos terminan entre los 10 y los 12 años, pero la reestructuración de estos ganglios se produce a lo largo de la vida de acuerdo con las condiciones cambiantes de la existencia. La involución de los ganglios linfáticos relacionada con la edad se puede observar desde la adolescencia. A medida que aumenta la edad, su número disminuye y el tamaño promedio de los nodos restantes aumenta. Esto se debe a que muchos ganglios linfáticos pequeños son reemplazados por completo por tejido adiposo y conectivo fibroso denso. Los nodos cercanos crecen juntos y forman nodos más grandes de forma segmentaria o similar a una cinta. A partir de la adolescencia aparecen grupos de células grasas en el estroma y parénquima de los ganglios linfáticos. A medida que aumenta la edad, aumenta la cantidad de tejido conectivo en los ganglios, la cápsula se engrosa y las trabéculas hiliares crecen notablemente; las fibras del estroma reticular se espesan. El tejido conectivo fibroso denso y el tejido adiposo con el aumento de la edad desplazan gradualmente el parénquima linfoide en los ganglios linfáticos. Al mismo tiempo, la cantidad de sustancia cortical en los ganglios disminuye y el contenido de la médula aumenta relativamente. Los senos intermedios en la médula se expanden. También hay características relacionadas con la edad de la composición celular de los ganglios linfáticos; aumenta el número de linfocitos pequeños y macrófagos, mientras que disminuye el número de linfocitos medianos.