Organske kiseline, lipidi, vitamini. Osnove biohemije lipida u ljudskom tijelu
Spiralizirani (organizirani) dijelovi nukleinskih kiselina su optički aktivni, rotiraju ravan polarizirane svjetlosti, apsorbiraju ultraljubičasto na maksimalno 260 nm, ali je intenzitet apsorpcije nukleinskih kiselina manji nego kod mješavine nukleotida - hipohromni efekat .
^ Predavanje #3 (2 sata)
Tema: Biohemija ugljenih hidrata
Prije stotinjak godina predloženo je da se ugljikohidrati nazivaju spojevima čiji sastav odgovara formuli (CHOH) n, odnosno ugljičnim hidratima. Štaviše, n, odnosno broj atoma ugljika, ovisno o vrsti ugljikohidrata, može varirati od tri do nekoliko hiljada. Biološke funkcije ugljikohidrata su raznolike i zavise od vrste ugljikohidrata ili njihovih derivata. Ugljikohidrati čine oko 2% mase životinjskog tijela (u biljkama ugljikohidrati čine do 80%). Najveća količina ugljikohidrata nalazi se u jetri, u vezivnom tkivu. Glavne funkcije ugljikohidrata su:
Energija (prilikom cijepanja 1 g ugljikohidrata formira se 4,1 - 4,2 kcal;
Plastični ili strukturni (građevinski);
Zaštitne (u kombinaciji s proteinima);
Regulatorni (također u kombinaciji s proteinima);
Receptor (takođe u kompleksu sa proteinima) itd.
Svi ugljikohidrati su podijeljeni u sljedeće klase:
1. Monosaharidi (monomerne jedinice), ponekad se nazivaju jednostavnim šećerima. Općenito, "saharid" - u prijevodu s grčkog - "slatko". I zaista, mnogi jednostavni šećeri imaju slatki ukus.
2. Oligosaharidi (dva ili više monomera povezanih u jedan lanac).
3. Polisaharidi - jedinjenja velike molekularne težine, koja sadrže od sto do nekoliko hiljada monomera.
Monosaharidi
Fisher je prvi put utvrdio strukturu nekih monosaharida u drugoj polovini 19. stoljeća. Svi jednostavni monosaharidi imaju opću empirijsku formulu (CHOH) n, gdje je n cijeli broj od tri do devet. Bez obzira na broj atoma ugljika, svi saharidi se mogu svrstati u jednu od dvije klase: aldoze ili ketoze (završetak -oza označava pripadnost ugljikohidratima). Aldoze sadrže aldehidnu funkcionalnu grupu, dok ketoze sadrže funkcionalnu grupu. Aldo- i keto-trioze, tetroze, pentoze itd. razlikuju se po broju atoma ugljika. Podjela na aldoze i ketoze može se izvršiti i po drugoj osnovi: ako se karbonilna grupa (= C =) nalazi na kraju lanca, onda je to aldoza, a ako se karbonilna grupa nalazi negdje drugdje, onda ovo je ketoza, odnosno aldehid ili keton.
^
Životinjska tkiva sadrže sljedeće monosaharide:
trioses

U prirodi prevladavaju šećeri s D-konfiguracijom:
tetroze

Broj izomera za monosaharide je 2 m, gdje je m broj asimetričnih atoma ugljika. U triozama je to C 2, u tetrozama - C 2, C 3, u pentozama - C 2, C 3, C 4, au hektozama - C 2, C 3, C 4, C 5 \u003d 16 izomera.
pentoze

![]()
heksoze


heptoze

Svi navedeni monošećeri ili ulaze u organizam hranom ili nastaju u metaboličkim procesima. Oni su bezbojne, kristalne čvrste supstance, veoma rastvorljive u vodi. Monošećer daje većinu energije i većinu ugljika potrebnih za sintezu proteina, masti i drugih ugljikohidrata.
U pravilu, monosaharidi s malim brojem ugljikovih atoma (trioze, tetroze) imaju strukturu u obliku ravnih lanaca. A monosaharidi s brojem ugljikovih atoma 5 i više imaju zatvorenu cikličku strukturu; da napišete ovu strukturu, koristite Haworthove formule (1929)

Zbog sličnosti sa organskom supstancom piranom  Glukoza napisana pomoću Heorsove formule naziva se -D-glukopironaza. Sve što se nalazi desno od atoma ugljika prilikom pisanja Fisherove formule nalazi se ispod kada se piše Hehorsova formula. i su određeni položajem grupe - OH na prvom atomu ugljika (desno ili dolje - ; lijevo ili gore - ). Posljednja CH 2 OH grupa (kod posljednjeg C atoma) je uvijek napisana na vrhu.
Glukoza napisana pomoću Heorsove formule naziva se -D-glukopironaza. Sve što se nalazi desno od atoma ugljika prilikom pisanja Fisherove formule nalazi se ispod kada se piše Hehorsova formula. i su određeni položajem grupe - OH na prvom atomu ugljika (desno ili dolje - ; lijevo ili gore - ). Posljednja CH 2 OH grupa (kod posljednjeg C atoma) je uvijek napisana na vrhu.
Petočlane prstenove karakteriše sličnost sa hemijskim jedinjenjem furan
 ,
stoga se monosaharidi napisani u ovom obliku nazivaju furanoze:
,
stoga se monosaharidi napisani u ovom obliku nazivaju furanoze:

Osim monosaharida, tjelesna tkiva sadrže i njihove derivate, koji nastaju u reakciji šećera pomoću reaktivnih grupa:
OH, - C \u003d O, \u003d C \u003d O, tj. alkohol, aldehid i keton.
glikozidi
Monosaharidi su sposobni da formiraju glikozidnu vezu tipa C-O-R. Najčešće, glikozidnu vezu formira alkoholna grupa (-OH) na prvom atomu ugljenika, iako druge grupe (-OH) mogu formirati glikozidnu vezu pod strožim uslovima. Na primjer:

To je, u stvari, reakcija dva alkohola koji formiraju jednostavan etar R–O–R. Uz pomoć glikozidne veze, monosaharidi se mogu međusobno povezati:

Oligosaharidi
Oligosaharidi uključuju šećere koji se sastoje od 2, 3, 4 ili više monosaharida. Laktoza se nalazi u tkivima (mliječni šećer - u majčinom mlijeku je oko 7%). Ovo je mješoviti oligosaharid (odnosno, sastoji se od različitih monosaharidnih ostataka), točnije, disaharida i sastoji se od ostataka glukoze i galaktoze povezanih α-glukoznom vezom:

Mješoviti oligosaharidi se nalaze u slobodnom obliku (u majčinom mlijeku) i povezani su s proteinima i lipidima (u tkivima). Uključuju glukozu, galaktozu, manozu, amino šećere, njihove acetil derivate, neuraminsku i sijaličnu kiselinu, L-fruktozu - ovo proizvodi tri-, tetra, - pentasaharide, itd. Prisustvo oligosaharidnih grupa u krvi i tkivima određuje specifičnost grupe ( krvne grupe), uzrokuju specifične međućelijske interakcije. Maltoza, saharoza i drugi oligosaharidi nalaze se u biljkama.
Bifidus faktor također spada u miješane oligosaharide, uključuje galaktozu, L-fukozu, N-acetilglukozamin, glukozu i neuraminsku kiselinu. Ovaj faktor je najvažniji supstrat za fermentacijsku mikrofloru.
Polisaharidi
Svi polisaharidi se mogu podijeliti na:
1. homopolisaharidi ( svi ostaci sastavnih monomera su identični). Jedini predstavnik u životinjskim tkivima je glikogen. Glikogen služi kao rezervni energetski materijal za ćeliju. Sastoji se od ostataka glukoze međusobno povezanih -1,4-glukozidnom vezom. Sve zajedno se odvija uz pomoć -1,6-glikozidne veze.

Grananje se dešava kroz 8-10 ostataka glukoze u glavnom lancu. Grananje glikogenskog lanca povećava njegovu topljivost, a također povećava brzinu sinteze i razgradnje glikogena, jer se povećava površina kontakta glikogena sa enzimima (glikogen sintazom i glikogen fosforilazom).
Glikogen je prisutan u svim ćelijama, ali najviše u jetri (5-10% ukupne mase) iu mišićima (1-2%). Molekularna težina glikogena je do nekoliko miliona. To je depo glukoze u tijelu.
2. Heteropolisaharidi(sastoje se od različitih monomera). Heteropolisaharidi su gotovo uvijek u kompleksu sa jednom ili drugom količinom proteina (od 1 do 5%), a kompleksi se nazivaju i proteoglikani. Saharidni dio takvog kompleksa naziva se mukopolisaharidi ili glikozaminoglikani. Mnogo je proteoglikana u vezivnom tkivu (do 33%), posebno u labavom vezivnom tkivu (međućelijska tvar), tetivama, ligamentima, koži, rožnjači, staklastom tijelu, srčanim zaliscima itd.
^ Varenje ugljikohidrata
Ovo je proces hidrolitičkog cijepanja poli- i oligosaharida. Probava počinje u ustima. Pljuvačka sadrži enzim koji razgrađuje škrob (-amilazu), razgrađuje -1,4-glikozidne veze unutar molekula (ali ne i terminalne), dok stvara dekstrine, malu količinu maltoze i izomaltoze. U želucu se ugljikohidrati ne probavljaju, jer. pH je kisel. Potpuna probava se događa u tankom crijevu. Gušterača proizvodi , -amilaze i otpušta ih u lumen crijeva. Ćelije crijevne sluznice također proizvode brojne enzime koji probavljaju oligosaharide. Jedna grupa enzima cijepa -glikozidne veze (maltaza: -1,4-glikozidne veze, izomaltaza--1,6; saharoza - saharoza). Druga grupa cijepa -glikozidne veze (laktaza, -galaktozidaza, heterogalaktozidaza - cijepa miješane oligosaharide). Pod djelovanjem ovih enzima dolazi do cijepanja na monosaharide. Probava je češće parijetalna (blizu površine ćelije). Neprobavlja se: vlakna (bez enzima), ali su potrebna za crijevnu pokretljivost, pektozani (polisaharidi koji se sastoje od pentoza). Tokom probave nastaje najveći dio glukoze. Apsorpcija se vrši uz pomoć posebnih nosača, dok je specifičnost karakteristična, energija se troši (povećava se potrošnja kisika, troši se ATP). Glukoza i galaktoza se mogu apsorbirati zbog aktivnog transporta, zbog gradijenta koncentracije Na + jona koje stvara Na, K - ATPaza. To osigurava njihovu apsorpciju čak i pri niskim koncentracijama u crijevima. Protokom krvi kroz portalnu venu glukoza se isporučuje u jetru, a djelomično se krvotokom doprema do svih organa i tkiva.
Glukoza može doći iz gastrointestinalnog trakta sa krvotokom, nastati prilikom razgradnje glikogena, a može se sintetizirati iz neugljikohidratnih komponenti - glukoneogenezom.
glikoliza
Glikoliza se općenito odnosi na razgradnju ugljikohidrata. U zavisnosti od specifičnog organizma i (ili) uslova njegovog rasta, glikoliza može ići anaerobnim putem (u ovom slučaju to se naziva anaerobna glikoliza).
I može pratiti aerobni put (odnosno, aerobnu glikolizu). Za mnoge mikroorganizme koji rastu u anaerobnim uvjetima, glikoliza je glavni katabolički put za dobivanje energije iz ugljikohidratnih supstrata, što dovodi do stvaranja određenih metaboličkih krajnjih proizvoda, na primjer, etanola, laktata, glicerola. Ova vrsta procesa se često naziva fermentacija. Kod životinja, anaerobna glikoliza (sa stvaranjem laktata) daje energiju za kontrakciju skeletnih mišića u uvjetima ograničene opskrbe kisikom. U nekim slučajevima, glikoliza (aerobna glikoliza) slijedi anaerobni put u aerobnim uvjetima, tj. krajnji proizvod je laktat, iako je kisik prisutan. Takvi se procesi odvijaju, na primjer, u eritrocitima, retini, crijevnoj sluznici, u tkivima fetusa (ubrzo nakon rođenja). Glikoliza je jedini anaerobni put koji osigurava energiju. U aerobnim uslovima (u prisustvu kiseonika), reakcije glikolize predstavljaju početni oblik razgradnje ugljenih hidrata, koji je dalje povezan sa ciklusom limunske kiseline. U tom slučaju, glikoliza se zaustavlja u fazi formiranja PVC-a. U aerobnim uvjetima, glikoliza i Krebsov ciklus zajedno dovode do potpune oksidacije heksoze u ugljični dioksid. Ovaj proces je praćen oslobađanjem velikih količina potencijalno dostupne metaboličke energije, uglavnom u obliku visokoenergetskih proizvoda NADH i FADH 2, koji, oksidirani tokom respiratornog lanca, daju energiju potrebnu za RP proces, što rezultira formiranje ATP-a. Glikoliza je prva potpuno dešifrovana sekvenca biologije hemijske reakcije, trebalo je skoro 50 godina (1890-1940). Za to vrijeme otkriveni su fosfoesteri, ATP, NAD, izolovani i okarakterisani mnogi faktori itd. Glikoliza se odvija u citosolu.
Obrazovanje je veoma važno završna faza NAD+, koji se koristi za oksidativnu fosforilaciju 3-fosfogliceraldehida.
Laktat je krajnji proizvod glikolize. Pošto laktat (kao i piruvat, inače) prilično lako prolazi kroz plazma membranu, iz mišićne ćelije (ili eritrocita) ulazi u krv, protokom krvi ulazi u jetru, gdje se oksidira u piruvat, koji zatim se pretvara u glukozu. Tako se formira svojevrsni ciklus. Ove transformacije čine Corey ciklus:

Generalno, tokom glikolize se formiraju 4 ATP molekula, a 2 ATP se troše. Dakle, ukupan efekat oksidacije jednog molekula glukoze tokom glikolize u laktat je 2 ATP. Iskoristi se samo oko 3% sve moguće energije, koja bi se mogla dobiti oksidacijom glukoze, tj. 27 kg glukoze bi se dnevno moralo razgraditi da bi se organizam opskrbio energijom (ako bi se energija dobivala samo glikolizom). Međutim, glikoliza se odvija u ćelijama. Potreban je u mišićima za brzi prijem energije.
^ Predavanje #4 (2 sata)
Tema: Biohemija lipida
Lipidi su prirodni nepolarni spojevi koji su potpuno ili gotovo potpuno netopivi u vodi, ali topljivi u nepolarnim otapalima, kao što su kloroform, ugljični disulfid, eter i vrući etanol. Glavna karakteristika ove grupe prilično heterogenih supstanci je njihova nepolarnost, što se u nekim slučajevima objašnjava prisutnošću u ovim spojevima jedne ili više masnih kiselina koje sadrže duge alifatske ugljikovodične lance. Sadržaj lipida u tijelu je 10 - 20% tjelesne težine.
Lipidi obavljaju sljedeće biološke funkcije :
1. One su komponente membrana koje u velikoj mjeri određuju njihovu propusnost.
2. Služe kao jedan od glavnih oblika skladištenja ugljenika i energije (1 g masti tokom oksidacije daje 9,3 kcal).
3. Mogu biti prekursori drugih važnih jedinjenja.
4. Djeluju kao zaštitne barijere koje štite od toplinskih, električnih i fizičkih utjecaja.
5. Oni su dio zaštitnih ljuski koje štite od infekcije i prekomjernog gubitka ili nakupljanja vode. Masti su izvori endogene vode.
6. U nekim slučajevima to su vitamini i hormoni.
7. Učestvuju u adukciji nervnih impulsa, jer su deo mijelinskih ovojnica.
Postoji klasifikacija zasnovana na sadržaju jednog ili drugog alkohola u sastavu masti. Shodno tome, izolovani su lipidi i holesterol koji sadrže glicerin, sfingozin i njegovi derivati.
Klasifikacija lipida prema njihovoj strukturi:
1. Jednostavni lipidi.
2. Kompleksni lipidi.
3. Lipidni derivati.
Jednostavne masti uključuju samo estre masnih kiselina i alkohola. Sastav složenih lipida, pored masnih kiselina i alkohola, uključuje i druge komponente. Među složenim lipidima razlikuju se fosfoacilgliceroli, sfingomijelini, cerebrozidi i gangliozidi. Derivati lipida uključuju sva jedinjenja koja se ne mogu jasno klasifikovati kao jednostavne ili složene lipide, kao što su steroidi, karotenoidi i vitamini lipidne prirode. Mnogi od njih nastaju od ugljikovodičnih lanaca koji su bili dio masnih kiselina.Sastav jednostavnih i složenih lipida nužno uključuje masne kiseline. U stvari, to su karboksilne kiseline sa dugim alifatskim lancem. Prirodne masne kiseline su veoma raznovrsne.
Radi praktičnosti, razlikuju se sljedeće grupe:
1. Većina masnih kiselina su monokarboksilne kiseline koje sadrže linearne ugljikovodične lance s parnim brojem atoma (obično C 12 - C 20). Manje uobičajene su FA sa kraćim lancima ili s neparnim brojem atoma ugljika.
2. Često postoje kiseline koje sadrže etilenske (nezasićene, dvostruke) veze (obično C 18 - C 20 kiseline). Ako postoje dvije ili više dvostrukih veza, one su gotovo uvijek odvojene jednom metilenskom grupom: - CH = CH-CH 2 -CH = CH -.
3. U nezasićenim kiselinama sa dvostrukim vezama, gotovo uvijek se odvija cis konfiguracija.
Veliki broj nepolarnih C-C i C-H veza u ugljičnom lancu daje suštinski nepolarni karakter cijeloj molekuli u cjelini, iako ima i polarnu grupu - COOH. Sasvim je jasno da će spoj koji sadrži jedan ili više FA ostataka biti nepolaran, što je razlog netopivosti lipida u vodi, a također uzrokuje sklapanje lipida u biomembrane.
Razmislite različite vrste lipidi:
^ 1. Jednostavni lipidi . Dijele se u dvije grupe: neutralni acilgliceroli i voskovi.
Neutralni acilgliceroli. U zavisnosti od broja FA ostataka vezanih za trihidrični alkohol glicerol, razlikuju se mono-, di-, triacil-gliceroli. Triacil derivati su najčešći u prirodi. U svim slučajevima, prosti acilglicerol ne sadrži funkcionalne jonske grupe i pripada neutralnim lipidima. Acil bočni lanci su obično različiti.

Voskovi. To su estri, međutim i alkohol (do C 18) i kiselina uključena u njihov sastav imaju duge ugljikovodične lance. Svi voskovi su potpuno nerastvorljivi u vodi. Prirodni voskovi su obično krajnji produkti onih metaboličkih puteva čija je glavna uloga stvaranje zaštitnih premaza. Ptičje perje i životinjska koža imaju voštani premaz koji ih čini vodoodbojnim. Voštani premaz listova i plodova biljaka smanjuje gubitak vlage i smanjuje mogućnost infekcije.
^ 2. Kompleksni lipidi.
Postoje tri glavne klase složenih lipida:
1. Fosfoacilgliceroli.
2. Sfingomijelini.
3. Glikolipidi.
Prve dvije klase nazivaju se fosfatidi ili fosfolipidi zbog prisustva fosfatnih grupa. Sva ova jedinjenja nalaze se samo u membranama, jer su njihove glavne komponente.
^ Fosfoacilgliceroli (PAG). Ovo su najčešći složeni lipidi. Broj različitih FA i njihova relativna koncentracija uvelike varira u zavisnosti od vrste ćelija i njihovog fiziološkog stanja. Uprkos ovoj raznolikosti, oni i dalje razlikuju glavne i sporedne komponente. Glavni: fosfatilholin (lecitin), fosfatidiletanolamin (cefalin), fosfatidilglicerin. manji: fosfatidilserin, difosfatidilglicerol (kardiolipid), fosfatidilinozitol i fosfatidna kiselina. Svi ostali FAG se formiraju od fosfatidne kiseline.
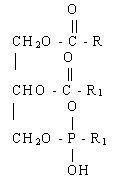
Fosfatna kiselina
Različiti FAG-ovi se međusobno razlikuju po dodatnim grupama vezanim fosfoesterskom vezom za fosfatidnu kiselinu. Sastav FA različitih FAG-ova se razlikuje čak i unutar istog organizma.
^ Struktura nekih FA:


U ovim strukturama se jasno vidi da jedinjenja imaju dva različita regiona: nepolarni hidrofobni „rep“ i polarnu hidrofilnu „glavu“. Takva dvojna priroda naziva se alifatska (od grčkog - dvostrano i osjećanje).
^ Sfingomijelini (fosfosvingolipidi). Sastoje se od jedne molekule masnih kiselina, holina, fosforne kiseline i swingozin alkohola. Sfingozini su porodica dugih nezasićenih lanaca amino alkohola koji se razlikuju po dužini lanca. FA je vezan za amino grupu sfingozina preko amidne veze i nastaje ceramid. Dodavanje ostatka fosforilholina dovršava formiranje swingomijelina:

^ Glikolipidi (glikosfingolipidi). Takođe su na bazi ceramida. Postoje cerebrozidi i gangliozidi. Još jedna sekvencijalna reakcija biosinteze lipida je enzimska konverzija prekursora ceramida u cerebrozide i gangliozide vezivanjem grupe ugljikohidrata na ceramid. Cerebrozidi i gangliozidi se uglavnom (iako ne isključivo) nalaze u ćelijskim membranama nervnog i moždanog tkiva i takođe pokazuju amfipatska svojstva. Protozoa cerebrozidi sadrže monosaharidnu grupu vezanu glikozidnom vezom za krajnju -OH grupu ceramida. U pravilu je to glukoza ili galaktoza.

^ 3. Lipidni derivati.
Ovo je veoma heterogena grupa jedinjenja. Njihova sličnost je ograničena samo slabom rastvorljivošću u vodi. Najvažniji članovi ove grupe su steroidi, karotenoidi i srodni vitamini lipidne prirode.
Steroidi. Nalaze se u svim organizmima gdje obavljaju različite funkcije. Kod ljudi igraju ulogu polnih hormona, emulgatora u varenju lipida i uključeni su u transport lipida kroz membrane. U krvnoj plazmi djeluju kao protuupalni agensi i kao regulatori nekih metaboličkih procesa. Svi steroidi imaju sličnu strukturu, koja se zasniva na perhidrociklopentanofenantrenu. Raznolikost steroidnih struktura je posljedica različitim stepenima nezasićenost i prisustvo nekoliko grupa na različitim pozicijama ciklusa.

Prisustvo bočnog lanca ugljikovodika (C 8 - C 10) na 17. poziciji i hidroksilne grupe na poziciji 3 karakteristično je za veliku grupu steroida zvanih steroli. Najvažniji i najčešći od njih je holesterol. Ulazi kao strukturna komponenta u ćelijske membrane, međutim njegova koncentracija u membranama varira (do 40% ukupnog sadržaja membranskih lipida). Kolesterol (zbog sadržaja konsolidirane strukture koja je manje fleksibilna od produženog lanca ugljikovodika) daje membrani veću krutost (snagu). Kolesterol je također primarni metabolički prekursor drugih važnih steroida, uključujući žučne kiseline i polne hormone. U nekim tkivima holesterol je prekursor vitamina D. Spolni steroidni hormoni: muški - testosteron (uzrokuje sve polne karakteristike) i ženski: estradiol i progesteron (uglavnom odgovorni za regulaciju menstrualnog ciklusa).

Prvo je izoliran iz žučnih kamenaca, pa otuda i naziv (chole - žuč, steros - tvrd).
Karotenoidi. Karotenoidi se dijele u dvije grupe: karoteni i ksantofili. Obje grupe uključuju pigmente netopive u vodi koji su široko rasprostranjeni u prirodi, a većina ih se nalazi u biljkama i algama. Karoteni su čisto ugljikovodična jedinjenja, dok ksantofili sadrže kisikove grupe. Karoteni su šire rasprostranjeni u prirodi. Najčešći je beta-karoten. To je C 40 ugljovodonik sa jako razgranatom strukturom i nezasićenim vezama, na oba kraja ugljovodoničnog lanca nalaze se identični supstituisani prstenovi. Svi ostali karotenoidi, u suštini, mogu se smatrati varijantama ove strukture. Tokom enzimskog cijepanja beta-karotena, dva molekula vitamina A nastaju na simetričan način.
^ Lipidni vitamini. vitamin D. Potreban za normalan metabolizam kalcijuma i fosfora neophodnih za rast zdravih kostiju i zuba. Njegov nedostatak uzrokuje rahitis - bolest u kojoj kosti postaju meke i plastične, što dovodi do njihove deformacije. Vitamin D nastaje iz prekursora sterola uz UV zračenje. Jedan važan prekursor je 7-dihidroholesterol, koji se i sam proizvodi enzimski iz holesterola. Iz njega se dobija vitamin D3. Postoji nekoliko oblika vitamina D. Oblik D 3 (kolikalceferol) je prisutan u mlijeku, puteru, ribljoj jetri, koji su glavni egzogeni izvor ovog vitamina. Vitamin D 3 može se sintetizirati u koži, pod uslovom da dobije normalan dio sunčeve svjetlosti.
vitamin E. Osnovna struktura vitamina E (a postoji nekoliko oblika) naziva se tokoferol. Najaktivniji oblik vitamina E je -tokoferol. Kod štakora je vitamin E neophodan za reprodukciju (ovo još nije dokazano za ljude). Kod nekih životinja nedostatak vitamina E dovodi do mišićne distrofije. Vitamin E se obično dodaje tokom obrade hrane jer sprečava kvarenje zbog svojih antioksidativnih svojstava. prehrambeni proizvodi zbog njihove oksidacije. U živim ćelijama vitamin E takođe deluje kao antioksidans, zajedno sa vitaminom C i glutationom.
vitamin K. Njegov nedostatak usporava zgrušavanje krvi. Njegova struktura je zasnovana na bicikličkom sistemu naftokinona, za koji je vezan dugačak ugljikovodični lanac. Može doći iz hrane (svježe povrće, sir), ili ga mogu stimulirati bakterije u gastrointestinalnom traktu. Njegov koagulacijski učinak je određen učešćem u sintezi protrombina, koji potom prelazi u trombin.
Biomembrane
Biomembrane zatvaraju ćeliju i mnoge organele unutar ćelije. Veoma je aktivan biološki sistemi, odgovorni za procese kao što su selektivni transport supstanci u i iz ćelije i subcelularnih odjeljaka (formacija), vezivanje hormona i drugih regulatornih molekula, reakcije katalizirane enzimima, prijenos električnih impulsa, pa čak i sinteza ATP-a . Membrane su različite i različite vrste membrane imaju različite tipove funkcionalna aktivnost. Na primjer, samo je unutrašnja mitohondrijska membrana odgovorna za biosintezu ATP-a.
^ Hemijski sastav. Membrane se sastoje od molekula lipida i proteina. Njihova relativna količina se značajno razlikuje za različite membrane, varirajući od 20% proteina + 80% lipida do 75% proteina + 25% lipida. Ugljikohidrati u obliku glikoproteina i glikolipida čine 0,5-10% membranske tvari.
^ Varenje lipida
Nedavno su se pojavili izvještaji o postojanju lipaze koju luči leđna površina jezika, a koja je uključena u probavu triacilglicerola u ishrani. Lipaza se također luči u želucu, ali zbog niskog pH ova lipaza je neophodna fiziološki značaj nema. Istovremeno, lipaza jezika može zadržati svoje djelovanje dva do četiri sata unutar bolusa hrane. Lipaza jezika je najaktivnija u odnosu na kratkolančane masne kiseline i pretežno djeluje na 3. poziciji. Posebno dobar supstrat za ovaj enzim je mlečna mast (uglavnom sadrži kratko- i srednjelančane masne kiseline, estrifikovane na 3. poziciji). Glavna probava lipida odvija se u tankom crijevu. Zapravo, probava masti se svodi na hidrolitičko cijepanje. Za to su potrebni sljedeći uslovi:
1. Prisustvo odgovarajućih enzima (lipaza);
2. Optium pH (5 - 7);
3. Masti moraju biti emulgirane (tj. zdrobljene), jer su u vodenoj sredini u obliku velikih kapi koje su nepristupačne djelovanju enzima, koji, budući da su topljivi u vodi, ne mogu prodrijeti u kap. Ako se ova kap drobi (emulgira), onda ukupna površina >> i, shodno tome >> djelovanje enzima. Lipaze cijepaju masne kiseline iz triacilglicerola na pozicijama 1 i 3. Najteži je dekolte na 2. poziciji. Ovaj proces katalizira enzim fosfolipaza A 2 . Dakle, nakon potpunog varenja, triacilgliceroli se razlažu na glicerol i masne kiseline. Ali, kao što je već spomenuto, za potpunu razgradnju masti u tankom crijevu, masti moraju biti emulgirane. U principu, emulzifikacija se svodi na:
1. Smanjena površinska napetost;
2. drobljenje;
3. Stabilizacija emulzije (prevlačenje malih kapljica masti proteinskom ljuskom). Važnu ulogu u emulzifikaciji masti imaju žučne kiseline koje nastaju iz holesterola.
^ Predavanje #5 (2 sata)
Tema: Strukturna organizacija ćelija i organskih sistema vezanih za ishranu
ćelijska struktura
Cell je osnovna elementarna jedinica živog objekta. Termin živi objekat općenito uključuje sve objekte sposobne za metabolizam I reprodukcija poput sebe.
Za održavanje ove dvije osnovne funkcije u procesu evolucije, u ćeliji su nastale određene strukture tzv ćelijske organele. Oni obezbeđuju koordiniran i kontrolisan tok glavnih reakcionih procesa neophodnih za stalnu manifestaciju vitalne funkcije.
Za postojanje živog organizma važne su sljedeće ćelijske organele: jezgra, mitohondrije, endoplazmatski retikulum, ribozomi, lizozomi i mikrotijela. Ćelijske membrane, ne samo da odvajaju živi organizam (ćeliju). okruženje, ali sudjeluju u formiranju određenih odjeljaka ćelije (funkcionalne podjele). Oni služe kao strukturni element svih ćelijskih organela i učestvuju u funkcionisanju većine njih. Masa membrane može doseći 80% mase ćelije. Nestrukturirana, koloidna masa koja ispunjava unutarćelijski prostor naziva se citosol.
Ponekad ćelija sadrži morfološki različite granule koje sadrže proizvode stanične aktivnosti, bilo proizvode skladištenja (glikogen, masne kapljice) ili proizvode koji se moraju transportovati iz ćelije (proenzimi).
Samostalno žive ćelije (jednoćelijski organizmi) obično sadrže sve gore navedene strukture, a osim toga imaju ćelijski zid i, u nekim slučajevima, kontraktilni aparat (cilije i flagele).
Kod višećelijskih organizama postoji razlika u funkcijama zasnovana na diferencijaciji struktura. Dakle, u diferenciranim stanicama viših organizama postoje razlike u broju ćelijskih organela (ponekad razlike u njihovoj finoj strukturi, na primjer, u broju krista u mitohondrijima ćelija jetre i srčanog mišića), kao i u njihovom različita distribucija unutar ćelije (na primjer, akumulacija mitohondrija u ćeliji gdje se odvijaju endergonski procesi).
Vanjski dio plazma membrane uključuje složene hemijske strukture (proteini, glikoproteini) i naziva se glikokaliks. Ove strukture služe da se međusobno prepoznaju po ćelijama date vrste i da prepoznaju ćelije drugih vrsta. Ako su ove strukture dio posebno diferenciranih stanica viših organizama, one služe kao antigeni i uzrokuju stvaranje različitih specifičnih antitijela.
Čak i u organima, ćelije obično nisu u bliskom kontaktu. To je zbog prisutnosti negativnih naboja na površini ćelije, koji se međusobno odbijaju. Kao rezultat, između stanica se formiraju uski prostori, čiji se zbir u cijelom organu ili organizmu označava kao međućelijski prostor. Slično, zbir svih komponenti unutar ćelije (npr. jezgro, mitohondrije, itd.) naziva se unutarćelijski prostor.
Sa funkcionalne tačke gledišta, ne mogu postojati živi organizmi (pa čak ni ćelije ili njihove organele) sa neograničenim vremenom svog postojanja.
Počevši od procesa diobe, sve ćelije prolaze kroz tzv životni ciklus, na čijem kraju ili dolazi do diobe sa pojavom nove ćelije, ili dolazi do smrti. Trajanje ovog ciklusa je specifično za vrstu i, prema modernim konceptima, varira od nekoliko sati do desetina godina. Tokom životni ciklusćelija prolazi kroz određene faze, koje traju različito vrijeme, ovisno o vrsti ćelije, a karakteriziraju je striktno specifični metabolički procesi. Ove faze su označene kao G 1, S, G 2 itd. Stanice koje se ne dijele dalje i umiru nakon određenog vremena (npr. stanice sive tvari mozga) su stalno u G 1 fazi. U ovu grupu ćelija spadaju takozvane diferencirane ćelije.
Činjenica da su neki od molekula DNK u jezgri povezani sa histonima ranije se smatrala molekularnom osnovom. diferencijaciju. Ovo gledište je odbačeno nakon što se pokazalo da histoni igraju odlučujuću ulogu u formiranju superheliksa, u čijim su fibrilima dio deoksiribonukleoproteina i oslobađaju se samo u procesu replikacije DNK.
U skladu sa savremenim konceptima, proces diferencijacije ćelija je kontinuiran, potpuno je nepovratan zbog funkcionalnog blokiranja gena. Pretpostavlja se da bi ovaj proces trebali kontrolirati proizvodi gena odgovornih za strukturu i vrijeme sinteze.
^ Opis tipične ćelije
Core, okružen dvostrukom membranom s porama, lokaliziran je u sredini ćelije. Tamne jezgre su vidljive unutar jezgara. Vanjska nuklearna membrana je dio endoplazmatskog retikuluma povezanog s Golgijev kompleks. Ribosomi se nalaze na površini endoplazmatski retikulum. Ovalne strukture okružene dvostrukom membranom čiji unutrašnji dio formira kriste mitohondrije. Lizozomi okružena jednim slojem membrane. Sadrže hidrolitičke enzime, od kojih je većina neaktivna kao proenzimi. Kod jednoćelijskih organizama odgovorni su za probavu tvari koje ulaze u ćeliju. U višim organizmima, lizozomi su uključeni u razgradnju stanica koje su prestale obavljati svoje funkcije. mikrozomi (peroksizomi)) su manji od lizosoma. Sadrže oksidaze koje kataliziraju oksidaciju spojeva koji su strani za ćeliju i stoga moraju biti uklonjeni iz nje (npr. lijekovi, aromatična jedinjenja, itd.). Ćelija okružena plazma membrana, koji je konstruisan tako da na određenim mestima postaje moguć direktan prenos jedinjenja iz ekstracelularnog prostora u jezgro. Prostor između organela ispunjen koloidnom suspenzijom bogatom proteinima (enzimima) naziva se citosol.

plazma membrana
Plazma membranu čine proteini (periferni i integralni) ugrađeni u lipidni dvosloj. ^ integralni proteini su glikoproteinske prirode. Njihov N-terminalni dio je dio unutrašnjeg fosfolipidnog sloja, u koji prodire dio peptidnog lanca bogat nepolarnim aminokiselinama (u spiralnoj konformaciji), a njihovi bočni lanci ulaze u brojne hidrofobne kontakte sa alifatskim fosfolipidnim lancima. Lanci oligosaharida, koji obično sadrže GlcNAc, Man, Cal, Fuc i N-acetilneuraminsku kiselinu, mogu biti povezani sa integralnim proteinskim peptidnim lancem na vanjskoj površini plazma membrane. N-acetilneuraminska kiselina obično stoji na kraju oligosaharidnog lanca i uzrokuje njegov negativni naboj. Oligosaharidi daju posebna svojstva ćelijskoj površini, omogućavajući joj da prepozna ćelije istog organa ili ćelije drugog tipa (antigenost, kontaktna inhibicija). Oligosaharidi formiraju sloj na površini ćelije koji se naziva glikokaliks. Strukture lokalizovane na površini ćelije sprečavaju bliski kontakt između ćelija. To dovodi do činjenice da se između ćelija pojavljuje manje-više uzak prostor ispunjen tekućinom. Uobičajeni naziv za takva mjesta u organu ili organizmu je međućelijski prostor. Zbir svih zapremina unutar ćelija naziva se intracelularni prostor.

Mitohondrije
Ovo je mjesto formiranja ATP-a. Energija potrebna za njegovu sintezu javlja se kao rezultat postupne oksidacije supstrata koji sadrže vodik (šećeri, lipidi, aminokiseline) u respiratornom lancu pod djelovanjem kisika. Dekarboksilacija u ciklusu limunske kiseline dovodi do stvaranja CO 2 , a rezultat oksidacije je stvaranje H 2 O. Enzimi koji obezbjeđuju prijenos elektrona dio su unutrašnje membrane mitohondrija. Kiseonik ulazi u mitohondrije difuzijom. Produkt mitohondrijalne aktivnosti (ATP) prenosi se usljed procesa translokacije sa mjesta nastanka u ekstramitohondrijski prostor, gdje se koristi. Kako bi se osigurao brz prijenos ATP-a, mitohondrije su lokalizirane u blizini struktura u kojima se odvijaju procesi koji troše energiju (na primjer, u blizini elemenata uključenih u proces kontrakcije).

Interfazno jezgro i hromozomi
Interfazno jezgro je ispunjeno supstancom koja se zove hromatin. Uz DNK, u hromatinu su prisutne dvije vrste proteina: glavni proteini su histoni i nehistonski proteini (koji su po pravilu kiseli). Hromatin se sastoji od ponavljajućih strukturnih elemenata - nukleozoma (ili v-tijela). Jezgro nukleosoma formiraju četiri tipa histona, dajući oktamer koji sadrži dva molekula svakog od histona H3, H4, H2A i H2B. Molekuli histona su međusobno povezani hidrofobnim interakcijama, a njihove N-terminalne sekvence (uglavnom pozitivno nabijene) nalaze se na površini oktamera. Ovo, zauzvrat, osigurava interakciju oktamera sa dvostrukom spiralom DNK. Ovako vezan fragment DNK sadrži, ovisno o biološkom objektu, 154-241 par baza. Udaljenost između dva ukleosoma je unutar 9-14 nm, a DNK koja se nalazi u ovoj regiji povezana je sa histonom H1. Genetske informacije sadržane u datom regionu DNK postaju dostupne tek nakon modifikacije molekula histona (na primjer, nakon njegove fosforilacije).
Molekuli DNK vezani za nukleozome se zatim umotaju u superkolu. Dakle, čak i udaljeni segmenti DNK mogu biti u neposrednoj blizini, formirajući, na primjer, diskontinuirane gene. Prije diobe stanice, kromatin je u ovom najkondenziranijem obliku i formira hromozome. Pretpostavlja se da se funkcija nehistonskih proteina manifestuje u procesu transkripcije.
Količina DNK u ćeliji je konstantna (6 pg u ćeliji sisara). Ova vrijednost odgovara 5,5 x 10 9 parova nukleotida. Molekularna težina DNK je 10 10 -10 11 . Dužina potpuno rastegnutog molekula morala bi biti nekoliko centimetara. U hromozomima, molekuli DNK su u visoko kondenzovanom obliku (44:1), a in vivo 7 10 -6 g DNK odgovara 1 mikronu.

Ribosomi, polizomi
Ribosomi i polizomi su sferni i nalaze se u citoplazmi ili slobodni ili vezani za membrane endoplazmatskog retikuluma. Ribosomi se sastoje od dvije podjedinice. Tokom sinteze proteina, mRNA se vezuje za malu podjedinicu. Nekoliko ribozoma (ne manje od 4 i ne više od 100) može se vezati za jedan mRNA molekul. Ovaj kompleks se naziva polizom (poliribosom). Ribosomi se mogu razbiti na podjedinice; ovaj proces zavisi od koncentracije magnezijevih jona. Svaka podjedinica je izgrađena od rRNA molekula i specifičnog skupa proteina. Broj ribozoma u bakterijskoj ćeliji dostiže 10 4 , u životinjskoj je oko 10 5 .

Endoplazmatski retikulum
Endoplazmatski retikulum je membranska struktura koja se nalazi u citoplazmi, blizu jezgra. U elektronskom mikroskopu vidljive su cijevi zvane cisterne, na čijoj vanjskoj površini se mogu locirati ribozomi. Stoga se ove strukture nazivaju granularnim ili grubim endoplazmatskim retikulumom (SER), za razliku od glatkog endoplazmatskog retikuluma (SER), koji nema povezane ribozome. Ribosomi su vezani za stranu citoplazme na kojoj se odvija sinteza proteina. Nakon završetka sinteze proteina, nastali peptidni lanci prolaze kroz membranu u cisternu i prenose se na određena mjesta u ćeliji ili u Golgijev kompleks. Prema sadašnjem gledištu, oni proteini koji se moraju ukloniti iz ćelije i, eventualno, proteinske komponente membrana, sintetiziraju se u grubom endoplazmatskom retikulumu, a proteini koje koristi stanica sintetiziraju se na slobodnim ribosomima.

Golgijev kompleks
Golgijev kompleks formiraju paralelni tubularni membranski sistemi koji su direktno povezani sa grubim endoplazmatskim retikulumom. specifični enzimi, glikoziltransferaze, koji kataliziraju vezivanje monosaharida za proteine pomoću glikozidnih veza (preko Ser ili Thr OH grupa, rjeđe kroz Asn amidne grupe), dio su membrana Golgijevog aparata. Monosaharidi su obično uključeni u ovaj proces u obliku derivata sa UDP ili CMP. Nakon spajanja ugljikohidratnog dijela, molekul proteina može napustiti ćeliju zbog procesa egzocitoza. Međutim, prije nego što se to dogodi, protein mora ostati u ćeliji određeno vrijeme. Stoga se sadržaj vakuola Golgijevog kompleksa postupno koncentrira (voda se eliminira) i proteinski proizvodi (čak i u kristalnom obliku) se talože u obliku granula.

Motorni aparat ćelije
I jednoćelijski i višećelijski organizmi mogu biti opremljeni cilijama i flagelama koje se nalaze na površini, što omogućava kretanje organizma ili njegove okoline. Ove strukture su građene od fibrilarnih proteina (sadrže globularne podjedinice), koji pod uticajem makroergijskih spojeva mogu promijeniti svoju prostornu strukturu (konformaciju). Ova promjena u konformaciji proteinske molekule je koordinirana i regulirana. Ultrastruktura cilija i flagela je ista kod svih eukariota. Sadrže devet obodnih parova vlakana i jedan centralni par. Svaki filament je izgrađen od dvanaest ili više protofibrila. Svaki protofibril je formiran od globularnih podjedinica prečnika oko 4,5 nm. Proteini su zvali dyneins, izolovani su od vanjskih niti. Oni pokazuju aktivnost ATPaze u prisustvu ATP-a i Mg 2+ i čini se da je promjena njihove konformacije važna za kretanje. Citoplazmatske mikrotubule imaju sličnu strukturu.
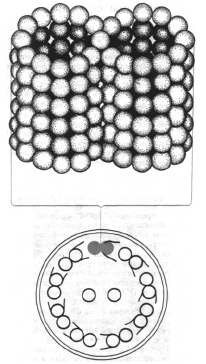
Rast i dioba ćelije
Podjelom matične ćelije nastaju dvije kćeri ćelije. U narednom periodu ćelije rastu i pripremaju se za dalju diobu. Interval između dvije mitoze ( ćelijski ciklus ) u fazi eksponencijalnog rasta je 10 minuta za bakterijsku ćeliju i 24 h za životinjsku. Za to vrijeme ćelija prolazi kroz nekoliko faza rasta. IN postmitotska faza G 1 ćelija sintetiše RNA molekule i proteine (DNK se ne sintetiše). Trajanje ove faze je 30-40% vremena cijelog ciklusa. Ćelije koje se dalje ne dijele (kao što su mišićne i glijalne ćelije) su stalno u G 1 fazi. U fazi sinteze (S) dolazi do potpunog umnožavanja DNK. U manjoj mjeri dolazi do sinteze RNK i proteina. Ova faza traje 30% vremena ciklusa. Priprema za mitozu se odvija u faza G 2 . Detalji ovog metaboličkog procesa trenutno su nepoznati, ali je jasno da se energija potrebna za mitozu u ovom trenutku pohranjuje. RNK i proteini nastavljaju da se sintetišu i tokom ovog perioda. Ova faza traje 10-20% vremena ciklusa. Mitoza zauzima 5-10% ciklusa, a za to vrijeme se ne dešavaju metabolički procesi.
U HeLa ćelijama, ukupno trajanje ciklusa je 23 h, G 1 =12 h, S=5 h, G 2 =5 h, mitoza =1 h.

BELORUSSKI DRŽAVNI UNIVERZITET ZA INFORMACIJU I RADIO ELEKTRONIKU
Odjel za ETT
"Osnove biohemije lipida u ljudskom tijelu"
MINSK, 2008
Lipidi su zajednički naziv za sve poznate masti i mastima slične supstance različite strukture, ali zajednička svojstva(nerastvorljivost u vodi, ekstrakcija neorganskim rastvaračima). Lipid (grčka mast).
U ljudskom tijelu, 10-20% masti od tjelesne težine. Lipidi su:
Protoplazmati - dio su svih ćelijskih struktura, organa i tkiva i praktično ostaju na istom nivou tokom cijelog života. Oni čine 25% ukupne tjelesne masti.
Rezervni lipidi se pohranjuju u tijelu, a njihova količina varira u zavisnosti od starosti, pola, uslova ishrane i aktivnosti.
Funkcije lipida u tijelu:
1 - plastična funkcija: učestvuju u izgradnji ćelijskih membrana svih organa i tkiva i formiranju mnogih biološki važnih jedinjenja (hormoni, vitamini rastvorljivi u mastima).
2 - energetska funkcija: lipidi obezbjeđuju 25-30% energetskih potreba tijela. Razgradnja 1 g masti je 9,3 kcal.
3 - masti su rezervni nutrijenti, njihov depo je potkožno tkivo, perirenalna kapsula.
4 – zaštitna funkcija lipidi: učestvuju u termoregulaciji, štite kožu od isušivanja, organe - od potresa mozga.
5- djeluju kao zaštitne membrane koje štite od infekcije ili pretjeranog gubitka ili nakupljanja vode.
6 - obezbeđuju apsorpciju vitamina rastvorljivih u mastima
Klasifikacija lipida:
1 - proste ili neutralne masti (estri masnih kiselina i alkohola).Neutralne masti se nalaze u organizmu ili u obliku protoplazmatske masti, koja je strukturna komponenta ćelija, ili u obliku rezervne, rezervne masti.
2 - kompleksne masti, su estri trihidričnog alkohola glicerola, masnih kiselina visoke molekularne težine i drugih komponenti. Među složenim mastima su: fosfolipidi, glikolipidi, sfignomijelini. Sfingolipidi se nalaze u membranama životinjskih i biljnih ćelija.
3 - derivati lipida. Ovo uključuje sva jedinjenja koja se ne mogu jasno klasifikovati kao jednostavne ili složene lipide, kao što su steroidi, karotenoidi i vitamini lipidne prirode.
4-voskovi - na primjer, lanolin, mješavina estera holesterola.
Voskovi su estri formirani od zasićenih i nezasićenih masnih kiselina i alkohola.
U neutralnim mastima nalaze se:
Masna kiselina.
Masne kiseline su dobile ime po načinu na koji su izolovane iz masti. To su karboksilne kiseline sa dugim alifatskim lancem.
Prirodne masne kiseline su veoma raznovrsne. Većina masnih kiselina su monokarboksilne kiseline koje sadrže linearne lance ugljikohidrata s parnim brojem atoma. Sadržaj nezasićenih masnih kiselina je veći od sadržaja zasićenih. Nezasićene masne kiseline imaju nižu tačku topljenja.
svojstva masnih kiselina.
Zasićene i nezasićene masne kiseline uvelike se razlikuju po svojoj strukturnoj konfiguraciji. U zasićenim masnim kiselinama, ugljikovodični rep može, u principu, poprimiti različite konformacije zbog potpune slobode rotacije oko terminalne jednostruke veze.
U nezasićenim kiselinama uočava se drugačija slika: nemogućnost rotacije oko dvostruke veze osigurava kruto savijanje ugljovodoničnog lanca.
Prirodne masne kiseline, zasićene i nezasićene, ne upijaju svjetlost ni u vidljivom ni u UV području. Spektrofotometrijski određeno tek nakon izomerizacije (230-260 nm). Nezasićene se određuju kvantitativnom titracijom. Analiza složenih smjesa masnih kiselina provodi se plinsko-tečnom hromatografijom.
Zasićene - palmitinske, stearinske, lipocerinske kiseline
Nezasićene: arahidonske, oleinske, linolne, linolenske.
Biljne masti se uglavnom sastoje od nezasićenih masnih kiselina.
Lipidi su obavezni sastavni dio uravnotežena ljudska ishrana. Odnos proteina, lipida i ugljenih hidrata treba da bude 1:1:4.
Vrijednost masti je veoma raznolika. Njihov visok sadržaj kalorija daje im posebnu vrijednost. Masti su rastvarači za vitamine A, D, E itd. Sa mastima se u organizam unose i neke nezasićene kiseline, koje se svrstavaju u esencijalne masne kiseline (linolna, linolenska, arahidonska), koje se ne sintetišu kod ljudi i životinja. Sa mastima u tijelo ulazi kompleks biološki aktivnih tvari: fosfolipidi, steroli.
Triacilgliceroli - njihova glavna funkcija je skladištenje lipida. Nalaze se u citosolu u obliku finih emulgiranih uljanih kapljica.
Kompleksne masti:
Fosfolipidi - glavne komponente ćelijskih membrana i subćelijskih organela, čine većinu tkiva mozga, nerava, jetre, srca, učestvuju u biosintezi proteina, aktivaciji protrombina, transportu lipida i vitamina rastvorljivih u mastima u krvi i limfi. Sastoje se od glicerola i dva molekula masnih kiselina, od kojih je jedan zasićen. a drugi je nezasićena + azotna baza.
Lipoproteini.
Polarni lipidi se povezuju s određenim specifičnim proteinima kako bi formirali lipoproteine, od kojih su transportni lipoproteini najpoznatiji i prisutni su u krvnoj plazmi sisara.
U takvim složenim lipidima, interakcije između lipida i proteinskih komponenti se odvijaju bez sudjelovanja kovalentnih veza.
Lipoproteini obično sadrže i polarne i neutralne lipide, kao i kolesterol i njegove estre. Oni služe kao oblik u kojem se lipidi transportuju iz tankog crijeva u jetru i iz jetre u masno tkivo i druga tkiva.
Nekoliko klasa lipoproteina je pronađeno u krvnoj plazmi, a njihova klasifikacija je zasnovana na razlikama u njihovoj gustoći. Lipoproteini s različitim omjerima lipida i proteina mogu se odvojiti u ultracentrifugi.
Najlakši lipoproteini su hilomikroni: velike strukture koje sadrže oko 80% triacilglicerola, 7% fosfoglicerida, 8% holesterola i njegovih estera i 2% proteina.
Beta-lipoproteini plazme sadrže 80-90% lipida, a alfa-lipoproteini - 40-70%.
Tačna struktura lipoproteina još uvijek nije poznata, ali postoje razlozi za vjerovanje da se proteinski lanac nalazi na vanjskoj površini, gdje formira tanku hidrofilnu ljusku oko micelarne lipidne strukture. Pohranjen u mastima ili trigliceridima večina energija koja se oslobađa kao rezultat hemijskih reakcija.
Pored nepolarnih lipida, postoje i polarni lipidi. One su glavne komponente ćelijskih membrana. Brojni enzimi i transportni sistemi su lokalizovani u membranama. Mnoga svojstva ćelijskih membrana su posljedica prisustva polarnih lipida u njima.
Membranski lipidi:
Membranski lipidi, zajedno sa ugljovodoničnim lancima, sadrže jednu ili više visoko polarnih „glava“. Fosfolipidi su prisutni u malim količinama u membranama. Njihova glavna komponenta - fosfogliceridi - sadrže 2 ostatka masnih kiselina koji esterificiraju prvu i drugu hidroksilnu grupu glicerola. Treća hidroksilna grupa formira estarsku vezu sa fosfornom kiselinom. Hidrolizira kada se zagrije s kiselinama i alkalijama, kao i enzimski - pod djelovanjem fosfolipaza.
Sfingolipidi su druga klasa membranskih lipida, imaju polarnu glavu i dva nepolarna repa, ali ne sadrže glicerol.
Podijeljeni su u 3 podklase: sfingomijelini, cerebrozidi i gangleozidi.
Sfingomijelini se nalaze u mijelinskim ovojnicama određenih tipova nervnih ćelija. Cerbrozidi - u membranama moždanih stanica. Gangleozidi su važne komponente specifičnih receptorskih mjesta smještenih na površini ćelijskih membrana. Nalaze se u onim specifičnim područjima nervnih završetaka u kojima dolazi do vezivanja molekula neurotransmitera u procesu hemijskog prenosa impulsa od jedne nervne ćelije do druge.
Eksterni ili plazma membrane proučavane su mnoge ćelije, kao i membrane brojnih intracelularnih organela, kao što su mitohondrije i hloroplasti. Sve membrane sadrže polarne lipide.
Lipidni dio membrane je mješavina polarnih lipida. Prirodne membrane karakteriše mala debljina (6-9nm) i elastičnost. Voda lako prolazi kroz membrane, ali su one praktički nepropusne za kontaminirane jone kao što su natrij, hlor ili vodonik i za polarne, ali ne kontaminirane molekule šećera. Polarni molekuli prodiru uz pomoć specifičnih nosača transportnog sistema.
Fosfogliceridi, sfingolipidi, glikolipidi i voskovi se često nazivaju lipidima koji se mogu saponificirati jer se sapuni formiraju kada se zagrijavaju (kao rezultat eliminacije masnih kiselina). Ćelije također sadrže manje lipida koji se ne saponiraju, ne hidroliziraju se uz oslobađanje masnih kiselina.
Postoje 2 vrste nesapunibilnih lipida:
Steroidi i terpeni
Steroidi - žučne kiseline, polni hormoni, hormoni nadbubrežne žlijezde.
Steroidi su široko rasprostranjeni u prirodi. Ova jedinjenja uključuju brojne supstance hormonalne prirode, kao i holesterol, žučne kiseline itd.
Steroli – holesterol Kolesterol igra ulogu međuproizvoda u sintezi mnogih drugih jedinjenja. Holesterol je bogat plazma membranama mnogih životinjskih ćelija, a nalazi se u mnogo manjim količinama u membranama mitohondrija i u endoplazmatskom retikulumu.
Biljke imaju fitosterole.
Terpeni - koji se nalaze u biljkama, mnogi od njih daju biljkama karakterističnu aromu i služe kao glavne komponente "mirisnih ulja".
vitamini rastvorljivi u mastima.
Vitamini su vitalne tvari prisutne u tijelu u tragovima i neophodne za normalne ćelijske funkcije. Vitamini rastvorljivi u mastima uključuju vitamine A, E, K, D, molekularna osnova njihovog delovanja nije tačno poznata.
Vitamini su klasifikovani kao lipidi jer su nerastvorljivi u vodi i mogu se ekstrahovati organskim rastvaračima. Vitamini rastvorljivi u mastima imaju izoprenoidnu strukturu (A, E, K), vitamin D je derivat steroida, iako steroidi potiču i od prekursora izoprenoida. Vitamin A se nalazi samo u životinjskim tkivima. Postoji u 2 hemijska oblika A-1 i A-2 (vitameri) - retinol 1 i retinol 2, i alkohol je koji sadrži aciklični prsten, za koji je vezan bočni lanac koji se sastoji od 2 jedinice izoprena.
Biljke sadrže karotenoidne pigmente. Alfa-, beta- i gama-karoteni, prilikom njihovog oksidativnog razgradnje u životinjskim tkivima, nastaju vitamin A. Nedostatak vitamina A dovodi do poremećaja rasta i razvoja „noćnog sljepila“, poremećena je normalna funkcija retinalnih štapića.
Vitamin E je predstavljen čitavom grupom vitamina prisutnih u biljnim uljima i naziva se tokoferoli. Ova jedinjenja imaju aromatični prstenasti sistem koji sadrži hidroksil i bočni lanac izoprenoida. Nedostatak dovodi do atrofije i slabosti mišića, steriliteta. Vjeruje se da ove tvari sprječavaju destruktivno djelovanje molekularnog kisika, ponekad se nazivaju i antioksidansima.
Vitamin K - K 1 i K 2, naftokinoni sa dugim bočnim izoprenoidnim lancima različite dužine. Nedostatak se očituje u kršenju procesa zgrušavanja krvi zbog gubitka sposobnosti tijela da sintetizira protrombin.
Vitamin D je derivat steroida. Najvažniji je vitamin D 2 - kalciferol i D 3. Nedostatak vitamina D dovodi osobu do poremećaja metabolizma kalcijuma i fosfora, što se manifestuje promjenama u strukturi kostiju i zuba. Vitamin D pospješuje apsorpciju Ca jona u tankom crijevu stimulirajući sintezu proteina uključenih u transport ovih jona.
Prostaglandini.
Ove tvari se nalaze u gotovo svim organima i tkivima ljudi i životinja, njihova visoka aktivnost, širok spektar djelovanja uporedivi su s djelovanjem hormona.
Prostaglandini su ciklične polinezasićene masne kiseline relativne molekulske težine 300-400. Sadrže samo ugljenik, kiseonik i vodonik.
Biosinteza prostaglandina se odvija u mikrosomima ćelije. Nezasićene masne kiseline su prekursori prostaglandina. Sintetiziraju se kao fiziološka potreba. Njihova uloga je najaktivniji nutrijenti. Smanjuju želučanu sekreciju, utiču na glatke mišiće, kardiovaskularni sistem.
Određuju se apsorpcionom spektrofotometrijom, gasno-tečnom hromatografijom, fluorescentnom analizom), kao i radioimunološkim metodama.
Soli žučnih kiselina.
Detergentni efekti žuči su posljedica žučnih soli. Steroidni dio molekule žučne kiseline ima hidrofobna svojstva karakteristična za lipide, a oksidirani bočni lanci su hidrofilni. Ova dvostruka rastvorljivost, karakteristična za deterdžente i sapune, označava se terminom amfipatičnost:
Hidrofobni kraj molekula se lako miješa s lipidima
Hidrofilni kraj se ne miješa lako s lipidima, ali olakšava kontakt sa vodenom fazom.
Ovo stvara emulziju lipida u vodi, koja se sastoji od sitnih kapljica masti.
Formiranje emulzija olakšava njihovu interakciju kao supstrata sa lipazama.
Druga vrsta je formiranje micela. U ovom slučaju, hidrofilni dijelovi stupaju u interakciju s vodenim medijem, stršeći prema van. Hidrofilni dijelovi ostaju međusobno povezani u unutrašnjoj sferi micele. Međutim, u miceli ne postoji ekstenzivna unutrašnja lipidna faza, jer njena debljina odgovara veličini jednog molekula. Micele su nekoliko redova veličine manje od čestica emulzije.
Emulzifikacija je važna jer povećava kontaktnu površinu kada lipaza djeluje na mišić, ali čestice emulzije su prevelike da prođu kroz staničnu membranu. Micele se lako apsorbuju u epitelnim ćelijama tankog creva.
Struktura ovih micela je takva da je njihovo hidrofobno jezgro (masne kiseline, gliceridi itd.) izvana okruženo hidrofilnom ljuskom masnih kiselina i fosfatida.
Kao dio micela, više masne kiseline i monogliceridi se prenose sa mjesta hidrolize masti na apsorbirajuću površinu ćelijskog epitela. Što se tiče mehanizma apsorpcije micela, ne postoji konsenzus:
1. Moguće, kao rezultat micelarne difuzije, a moguće i pinocitoze, micele kao cjelina čestica prodiru u epitelne ćelije resica. Ovdje dolazi do razgradnje micela masti, dok žučne kiseline odmah ulaze u krvotok i kroz sistem portalna vena u jetru, odakle se ponovo izlučuje u žuči.
2. Samo lipidna komponenta masnih micela može proći u ćelije resica, dok same žučne kiseline, ispunivši svoju fiziološku funkciju, ostaju u lumenu crijeva. I tek tada se apsorbiraju u krv (u ileumu), ulaze u jetru, a zatim se izlučuju žuči.
Trigliceridi i fosfolipidi koji se resintetiziraju u epitelnim stanicama crijeva, kao i kolesterol koji ulazi u ove stanice, spajaju se s malom količinom proteina i formiraju relativno stabilne kompleksne čestice - hilomikrone. Velike su veličine i ne mogu prodrijeti u krvne kapilare i difundirati u limfni sistem crijeva, a iz njega u torakalni limfni kanal i dalje u krvotok, tj. uz njihovu pomoć, endogeni trigliceridi, holesterol i delimično fosfolipidi se transportuju iz creva u krv.
Intracelularna hidroliza lipida.
Izvor masnih kiselina koje se podvrgavaju oksidaciji u tkivima viših životinja su ili ekstracelularna tečnost ili endogeni intracelularni lipidi. U mišićnom tkivu, uključujući srčani mišić, masne kiseline prolaze kroz direktnu oksidaciju. Glavni izvor endogene masne kiseline - rezervne masti sadržane u citoplazmi. Masne kiseline se prvo hidroliziraju specifičnim intracelularnim enzimima lipaze u glicerol i slobodne masne kiseline, a potonje se podvrgavaju aktivaciji i oksidaciji. Drugi izvor masnih kiselina su membranski fosfogliceridi. Metabolička obnova fosfoglicerida se stalno odvija, pri čemu nastaju slobodne masne kiseline.
Razgradnja masti.
Pljuvačka ne sadrži enzime za cijepanje masti. U želucu također nema primjetne probave masti iz hrane, ali dolazi do djelomične destrukcije lipoproteinskih kompleksa ćelijskih membrana hrane, što masti čini pristupačnijim za naknadno izlaganje lipazi soka pankreasa.
Razgradnja masti kod ljudi se događa uglavnom u gornjim dijelovima tankog crijeva, gdje postoje uslovi za emulgiranje masti. Najjače emulgirajuće dejstvo na masti imaju žučne soli koje sa žučom ulaze u duodenum u obliku soli natrijuma. Žučne kiseline su krajnji proizvod metabolizma holesterola. Po svojoj hemijskoj prirodi, žučne kiseline su derivati holanske kiseline. Žučne kiseline su prisutne u žuči u konjugiranom obliku. Ove veze se ponekad nazivaju uparene, jer. sastoje se od dvije komponente: žučne kiseline i glicina, te žučne kiseline i taurina. Žučne soli smanjuju površinsku napetost na površini vode i masti. Žučne kiseline takođe igraju važnu ulogu kao aktivator pankreasne lipaze, pod čijim uticajem dolazi do razgradnje masti u crevima. Lipaza koju proizvodi pankreas razgrađuje trigliceride koji su u emulgiranom stanju. Intestinalna lipaza je također uključena u razgradnju masti, katalizuje hidrolitičku razgradnju monoglicerida i ne djeluje na di- i trigliceride. Tako se u tankom crijevu glavni dio masti apsorbira nakon cijepanja na masne kiseline i glicerol, koji se, budući da su vrlo topljivi u vodi, apsorbiraju u crijevima, a zatim ulaze u krvotok portalne vene i odatle u jetra.
Dugolančane masne kiseline i monogliceridi se apsorbuju uz učešće žuči (žučne kiseline). Masne kiseline i monogliceridi formiraju micele koje su stabilne u vodenom mediju. Glavni endogeni izvor masnih kiselina je rezervna mast. Trigliceridi depoa masti igraju istu ulogu u metabolizmu lipida kao glikogen jetre u metabolizmu ugljikohidrata. Samo slobodne masne kiseline mogu se koristiti kao izvor energije, a trigliceridi moraju biti hidrolizirani lipazama u glicerol i slobodne masne kiseline. Mogu prijeći iz masnih depoa u krvnu plazmu i koristiti se kao energetski materijal.
Drugi izvor masnih kiselina su membranski fosfolipidi. Ćelije kontinuirano prolaze kroz metaboličku obnovu fosfolipida, pri čemu nastaju slobodne masne kiseline (fosfolipidi).
Oksidacija masnih kiselina u ćelijama odvija se u mitohondrijima uz učešće multienzimskog kompleksa. Proces oksidacije masnih kiselina sastoji se od sljedećih koraka:
Faza 1 - aktivacija masnih kiselina i njihov prodor iz citoplazme u mitohondrije, uz stvaranje acil-CoA. Uključuje 3 faze:
a) enzimska esterifikacija slobodne masne kiseline citoplazmatskim CoA (na račun ATP energije) u vanjskoj membrani mitohondrija.
b) transfer ostatka masne kiseline sa CoA na molekul karnitina, uz pomoć kojeg se ovaj ostatak transportuje kroz unutrašnju mitohondrijalnu membranu.
c) prijenos ostatka masne kiseline sa karnitina na intramitohondrijalni CoA.
Faza 2 - prva faza dehidrogenacije:. Kada se masne kiseline oksidiraju, to slijedi fazu aktivacije i događa se unutar mitohondrija. Acyl-CoA prolazi kroz enzimsku dehidrogenaciju, pretvarajući se u CoA ester nezasićene kiseline.
Faza 3 - faza hidratacije: dodavanje molekule vode i formiranje b-hidroksil CoA.
Faza 4 - druga faza dehidrogenacije, formira se b-ketoacil CoA.
Faza 5 - reakcija tiolaze: u njoj nastaje acetil CoA, koji se podvrgava oksidaciji u Krebsovom ciklusu, i acil CoA, koji zatim prolazi putem b oksidacije. Korak tiolitičkog cijepanja je tioliza, vrlo egzorganska reakcija.
Sekvencijalno oksidativno cijepanje molekula acetil CoA od CoA estera zasićenih masnih kiselina s dugim lancem naziva se b oksidacija.
Energetski bilans: U slučaju oksidacije, na primjer palmitinske kiseline, G je 9797 kJ.
LITERATURA
Komponente ćelija i njihovih membrana. Riboza i deoksiriboza, šećeri koji sadrže 5 atoma ugljika su dio ribonukleinske (RNA) i deoksiribonukleinske (DNK) kiselina. Metabolizam ugljenih hidrata u ljudskom organizmu sastoji se uglavnom od sledećih procesa: 1. Cepanje u gastrointestinalnom traktu na monosaharide, di- i polisaharide koji dolaze sa hranom. Apsorpcija u krv u crijevima. ...
Narandžasti krajnici i nakupljanje estera holesterola u drugim retikuloendotelnim tkivima. Patologija je povezana s ubrzanim katabolizmom apo A-I. Varenje i apsorpcija lipida. Bile. Značenje. U zoru formiranja moderne doktrine o egzokrinoj funkciji jetre, kada su prirodnjaci imali samo prve ...




Hanseleit je 1932. godine izveo jednadžbe za reakcije sinteze uree, koje su predstavljene u obliku ciklusa, koji se u literaturi naziva ornitinskim ciklusom Krebsove formacije uree. Treba istaći da je u biohemiji ovo bio prvi ciklični metabolički sistem, čiji je opis skoro 5 godina prethodio otkriću drugog metaboličkog procesa G. Krebsa, ciklusa trikarboksilne kiseline. Dalje...





Interakcija ćelija Vanjski sloj ćelijskih membrana Steroidi Komponente membrane; prekursori u sintezi žučnih kiselina i steroidnih hormona Ćelijske membrane; lipoproteini u krvi 3. Uloga lipida u ishrani čovjeka Biljne masti i ulja su esencijalni sastojak hrane, izvor energije i plastičnog materijala za čovjeka, snabdjevač niza potrebnih za njega...
BELORUSSKI DRŽAVNI UNIVERZITET ZA INFORMACIJU I RADIO ELEKTRONIKU
Odjel za ETT
"Osnove biohemije lipida u ljudskom tijelu"
MINSK, 2008
Lipidi su zajednički naziv za sve poznate masti i mastima slične supstance različite strukture, ali zajedničkih svojstava (nerastvorljivost u vodi, ekstrakcija neorganskim rastvaračima). Lipid (grčka mast).
U ljudskom tijelu, 10-20% masti od tjelesne težine. Lipidi su:
Protoplazmatski- dio su svih struktura ćelija, organa i tkiva i praktično ostaju na istom nivou tokom cijelog života. Oni čine 25% ukupne tjelesne masti.
Rezervirajte lipide- deponuju se u organizmu, a njihov broj varira u zavisnosti od starosti, pola, uslova ishrane, vrste aktivnosti.
Funkcije lipida u tijelu :
1 – plastična funkcija: učestvuju u izgradnji ćelijskih membrana svih organa i tkiva i formiranju mnogih biološki važnih jedinjenja (hormoni, vitamini rastvorljivi u mastima).
2 – energetska funkcija: lipidi obezbjeđuju 25-30% energetskih potreba tijela. Razgradnja 1 g masti je 9,3 kcal.
3 - masti su rezervne hranljive materije, njihov depo - potkožno tkivo, perirenalna kapsula.
4 – zaštitni funkcija lipida: uključeni su u termoregulaciju, štite kožu od isušivanja, organe od potresa mozga.
5- obavljati funkciju zaštitne školjke, štiteći od infekcije ili prekomjernog gubitka ili nakupljanja vode.
6 - obezbeđuju apsorpciju vitamina rastvorljivih u mastima
Klasifikacija lipida:
1 - jednostavno ili neutralne masti (estri masnih kiselina i alkohola).Neutralne masti se u organizmu nalaze ili u obliku protoplazmatske masti, koja je strukturna komponenta ćelija, ili u obliku rezervne, rezervne masti.
2 - složene masti, su estri trihidričnog alkohola glicerola, masnih kiselina visoke molekularne težine i drugih komponenti. Među složenim mastima su: fosfolipidi, glikolipidi, sfignomijelini. Sfingolipidi se nalaze u membranama životinjskih i biljnih ćelija.
3 - derivati lipida. Ovo uključuje sva jedinjenja koja se ne mogu jasno klasifikovati kao jednostavne ili složene lipide, kao što su steroidi, karotenoidi i vitamini lipidne prirode.
4- vosak- na primjer, lanolin, mješavina estera holesterola.
vosak - To su estri formirani od zasićenih i nezasićenih masnih kiselina i alkohola.
Nalazi se u neutralnim mastima :
Masna kiselina.
Masne kiseline su dobile ime po načinu na koji su izolovane iz masti. To su karboksilne kiseline sa dugim alifatskim lancem.
Prirodne masne kiseline su veoma raznovrsne. Većina masnih kiselina su monokarboksilne kiseline koje sadrže linearne lance ugljikohidrata s parnim brojem atoma. Sadržaj nezasićenih masnih kiselina je veći od sadržaja zasićenih. Nezasićene masne kiseline imaju nižu tačku topljenja.
svojstva masnih kiselina.
Zasićene i nezasićene masne kiseline uvelike se razlikuju po svojoj strukturnoj konfiguraciji. U zasićenim masnim kiselinama, ugljikovodični rep može, u principu, poprimiti različite konformacije zbog potpune slobode rotacije oko terminalne jednostruke veze.
U nezasićenim kiselinama uočava se drugačija slika: nemogućnost rotacije oko dvostruke veze osigurava kruto savijanje ugljovodoničnog lanca.
Prirodne masne kiseline, zasićene i nezasićene, ne upijaju svjetlost ni u vidljivom ni u UV području. Spektrofotometrijski određeno tek nakon izomerizacije (230-260 nm). Nezasićene se određuju kvantitativnom titracijom. Analiza složenih smjesa masnih kiselina provodi se plinsko-tečnom hromatografijom.
Zasićen-palmitinska, stearinska, lipocerinska kiselina
Nezasićene: arahidonska, oleinska, linolna, linolenska.
Biljne masti se uglavnom sastoje od nezasićenih masnih kiselina.
Lipidi su bitan dio uravnotežene ljudske prehrane. Odnos proteina, lipida i ugljenih hidrata treba da bude 1:1:4.
Vrijednost masti je veoma raznolika. Njihov visok sadržaj kalorija daje im posebnu vrijednost. Masti su rastvarači za vitamine A, D, E itd. Sa mastima se u organizam unose i neke nezasićene kiseline, koje se svrstavaju u esencijalne masne kiseline (linolna, linolenska, arahidonska), koje se ne sintetišu kod ljudi i životinja. Sa mastima u tijelo ulazi kompleks biološki aktivnih tvari: fosfolipidi, steroli.
Triacilgliceroli Njihova glavna funkcija je skladištenje lipida. Nalaze se u citosolu u obliku finih emulgiranih uljanih kapljica.
Kompleksne masti :
Fosfolipidi- glavne komponente ćelijskih membrana i subćelijskih organela, čine većinu tkiva mozga, nerava, jetre, srca, učestvuju u biosintezi proteina, aktivaciji protrombina, transportu lipida i vitamina rastvorljivih u mastima u krvi i limfi. Sastoje se od glicerola i dva molekula masnih kiselina, od kojih je jedan zasićen. a drugi je nezasićena + azotna baza.
Lipoproteini .
Polarni lipidi se povezuju s određenim specifičnim proteinima kako bi formirali lipoproteine, od kojih su transportni lipoproteini najpoznatiji i prisutni su u krvnoj plazmi sisara.
U takvim složenim lipidima, interakcije između lipida i proteinskih komponenti se odvijaju bez sudjelovanja kovalentnih veza.
Lipoproteini obično sadrže i polarne i neutralne lipide, kao i kolesterol i njegove estre. Oni služe kao oblik u kojem se lipidi transportuju iz tankog crijeva u jetru i iz jetre u masno tkivo i druga tkiva.
Nekoliko klasa lipoproteina je pronađeno u krvnoj plazmi, a njihova klasifikacija je zasnovana na razlikama u njihovoj gustoći. Lipoproteini s različitim omjerima lipida i proteina mogu se odvojiti u ultracentrifugi.
Najlakši lipoproteini su hilomikroni: velike strukture koje sadrže oko 80% triacilglicerola, 7% fosfoglicerida, 8% holesterola i njegovih estera i 2% proteina.
Beta-lipoproteini plazme sadrže 80-90% lipida, a alfa-lipoproteini - 40-70%.
Tačna struktura lipoproteina još uvijek nije poznata, ali postoje razlozi za vjerovanje da se proteinski lanac nalazi na vanjskoj površini, gdje formira tanku hidrofilnu ljusku oko micelarne lipidne strukture. Masti ili trigliceridi pohranjuju većinu energije koja se oslobađa kao rezultat kemijskih reakcija.
Pored nepolarnih lipida, postoje i polarni lipidi. One su glavne komponente ćelijskih membrana. Brojni enzimi i transportni sistemi su lokalizovani u membranama. Mnoga svojstva ćelijskih membrana su posljedica prisustva polarnih lipida u njima.
Membranski lipidi:
Membranski lipidi, zajedno sa ugljovodoničnim lancima, sadrže jednu ili više visoko polarnih „glava“. Fosfolipidi su prisutni u malim količinama u membranama. Njihova glavna komponenta - fosfogliceridi - sadrže 2 ostatka masnih kiselina koji esterificiraju prvu i drugu hidroksilnu grupu glicerola. Treća hidroksilna grupa formira estarsku vezu sa fosfornom kiselinom. Hidrolizira kada se zagrije s kiselinama i alkalijama, kao i enzimski - pod djelovanjem fosfolipaza.
Sfingolipidi su druga klasa membranskih lipida, imaju polarnu glavu i dva nepolarna repa, ali ne sadrže glicerol.
Podijeljeni su u 3 podklase: sfingomijelini, cerebrozidi i gangleozidi.
Sfingomijelini se nalaze u mijelinskim ovojnicama određenih tipova nervnih ćelija. Cerbrozidi - u membranama moždanih stanica. Gangleozidi su važne komponente specifičnih receptorskih mjesta smještenih na površini ćelijskih membrana. Nalaze se u onim specifičnim područjima nervnih završetaka u kojima dolazi do vezivanja molekula neurotransmitera u procesu hemijskog prenosa impulsa od jedne nervne ćelije do druge.
Proučavane su vanjske ili plazma membrane mnogih stanica, kao i membrane brojnih intracelularnih organela, poput mitohondrija i hloroplasta. Sve membrane sadrže polarne lipide.
Lipidni dio membrane je mješavina polarnih lipida. Prirodne membrane karakteriše mala debljina (6-9nm) i elastičnost. Voda lako prolazi kroz membrane, ali su one praktički nepropusne za kontaminirane jone kao što su natrij, hlor ili vodonik i za polarne, ali ne kontaminirane molekule šećera. Polarni molekuli prodiru uz pomoć specifičnih nosača transportnog sistema.
Fosfogliceridi, sfingolipidi, glikolipidi i voskovi se često nazivaju lipidima koji se mogu saponificirati jer se sapuni formiraju kada se zagrijavaju (kao rezultat eliminacije masnih kiselina). Ćelije također sadrže manje lipida koji se ne saponiraju, ne hidroliziraju se uz oslobađanje masnih kiselina.
Postoje 2 vrste nesapunibilnih lipida:
Steroidi i terpeni
Steroidi - žučne kiseline, polni hormoni, hormoni nadbubrežne žlijezde.
Steroidi su široko rasprostranjeni u prirodi. Ova jedinjenja uključuju brojne supstance hormonalne prirode, kao i holesterol, žučne kiseline itd.
Steroli – holesterol Kolesterol igra ulogu međuproizvoda u sintezi mnogih drugih jedinjenja. Holesterol je bogat plazma membranama mnogih životinjskih ćelija, a nalazi se u mnogo manjim količinama u membranama mitohondrija i u endoplazmatskom retikulumu.
Biljke imaju fitosterole.
Terpeni - koji se nalaze u biljkama, mnogi od njih daju biljkama karakterističnu aromu i služe kao glavne komponente "mirisnih ulja".
vitamini rastvorljivi u mastima.
Vitamini su vitalne tvari prisutne u tijelu u tragovima i neophodne za normalne ćelijske funkcije. Vitamini rastvorljivi u mastima uključuju vitamine A, E, K, D, molekularna osnova njihovog delovanja nije tačno poznata.
Vitamini su klasifikovani kao lipidi jer su nerastvorljivi u vodi i mogu se ekstrahovati organskim rastvaračima. Vitamini rastvorljivi u mastima imaju izoprenoidnu strukturu (A, E, K), vitamin D je derivat steroida, iako steroidi potiču i od prekursora izoprenoida. Vitamin A se nalazi samo u životinjskim tkivima. Postoji u 2 hemijska oblika A-1 i A-2 (vitameri) - retinol 1 i retinol 2, i alkohol je koji sadrži aciklični prsten, za koji je vezan bočni lanac koji se sastoji od 2 jedinice izoprena.
Biljke sadrže karotenoidne pigmente. Alfa-, beta- i gama-karoteni, prilikom njihovog oksidativnog razgradnje u životinjskim tkivima, nastaju vitamin A. Nedostatak vitamina A dovodi do poremećaja rasta i razvoja „noćnog sljepila“, poremećena je normalna funkcija retinalnih štapića.
vitamin E Predstavljen je cijelom grupom vitamina prisutnih u biljnim uljima, a nazivaju se tokoferoli. Ova jedinjenja imaju aromatični prstenasti sistem koji sadrži hidroksil i bočni lanac izoprenoida. Nedostatak dovodi do atrofije i slabosti mišića, steriliteta. Vjeruje se da ove tvari sprječavaju destruktivno djelovanje molekularnog kisika, ponekad se nazivaju i antioksidansima.
vitamin K– K 1 i K 2, naftokinoni sa dugim bočnim izoprenoidnim lancima različitih dužina. Nedostatak se očituje u kršenju procesa zgrušavanja krvi zbog gubitka sposobnosti tijela da sintetizira protrombin.
vitamin D- derivat steroida. Najvažniji je vitamin D 2 - kalciferol i D 3. Nedostatak vitamina D dovodi osobu do poremećaja metabolizma kalcijuma i fosfora, što se manifestuje promjenama u strukturi kostiju i zuba. Vitamin D pospješuje apsorpciju Ca jona u tankom crijevu stimulirajući sintezu proteina uključenih u transport ovih jona.
Prostaglandini.
Ove tvari se nalaze u gotovo svim organima i tkivima ljudi i životinja, njihova visoka aktivnost, širok spektar djelovanja uporedivi su s djelovanjem hormona.
Prostaglandini su ciklične polinezasićene masne kiseline relativne molekulske težine 300-400. Sadrže samo ugljenik, kiseonik i vodonik.
Biosinteza prostaglandina se odvija u mikrosomima ćelije. Nezasićene masne kiseline su prekursori prostaglandina. Sintetiziraju se kao fiziološka potreba. Njihova uloga je najaktivniji nutrijenti. Smanjuju želučanu sekreciju, utiču na glatke mišiće, kardiovaskularni sistem.
Određuju se apsorpcionom spektrofotometrijom, gasno-tečnom hromatografijom, fluorescentnom analizom), kao i radioimunološkim metodama.
Soli žučnih kiselina.
Detergentni efekti žuči su posljedica žučnih soli. Steroidni dio molekule žučne kiseline ima hidrofobna svojstva karakteristična za lipide, a oksidirani bočni lanci su hidrofilni. Ova dvostruka rastvorljivost, karakteristična za deterdžente i sapune, označava se terminom amfipatičnost:
Hidrofobni kraj molekula se lako miješa s lipidima
Hidrofilni kraj se ne miješa lako s lipidima, ali olakšava kontakt sa vodenom fazom.
Ovo stvara emulziju lipida u vodi, koja se sastoji od sitnih kapljica masti.
Formiranje emulzija olakšava njihovu interakciju kao supstrata sa lipazama.
Druga vrsta je formiranje micela. U ovom slučaju, hidrofilni dijelovi stupaju u interakciju s vodenim medijem, stršeći prema van. Hidrofilni dijelovi ostaju međusobno povezani u unutrašnjoj sferi micele. Međutim, u miceli ne postoji ekstenzivna unutrašnja lipidna faza, jer njena debljina odgovara veličini jednog molekula. Micele su nekoliko redova veličine manje od čestica emulzije.
Emulzifikacija je važna jer povećava kontaktnu površinu kada lipaza djeluje na mišić, ali čestice emulzije su prevelike da prođu kroz staničnu membranu. Micele se lako apsorbuju u epitelnim ćelijama tankog creva.
Struktura ovih micela je takva da je njihovo hidrofobno jezgro (masne kiseline, gliceridi itd.) izvana okruženo hidrofilnom ljuskom masnih kiselina i fosfatida.
Kao dio micela, više masne kiseline i monogliceridi se prenose sa mjesta hidrolize masti na apsorbirajuću površinu ćelijskog epitela. Što se tiče mehanizma apsorpcije micela, ne postoji konsenzus:
1. Moguće, kao rezultat micelarne difuzije, a moguće i pinocitoze, micele kao cjelina čestica prodiru u epitelne ćelije resica. Tu se micele masti razgrađuju, dok žučne kiseline odmah ulaze u krvotok i kroz sistem portalne vene do jetre, odakle se ponovo luče kao dio žuči.
2. Samo lipidna komponenta masnih micela može proći u ćelije resica, dok same žučne kiseline, ispunivši svoju fiziološku funkciju, ostaju u lumenu crijeva. I tek tada se apsorbiraju u krv (u ileumu), ulaze u jetru, a zatim se izlučuju žuči.
Trigliceridi i fosfolipidi koji se resintetiziraju u epitelnim stanicama crijeva, kao i kolesterol koji ulazi u ove stanice, spajaju se s malom količinom proteina i formiraju relativno stabilne kompleksne čestice - hilomikrone. Velike su veličine i ne mogu prodrijeti u krvne kapilare i difundirati u limfni sistem crijeva, a iz njega u torakalni limfni kanal i dalje u krvotok, tj. uz njihovu pomoć, endogeni trigliceridi, holesterol i delimično fosfolipidi se transportuju iz creva u krv.
Intracelularna hidroliza lipida .
Izvor masnih kiselina koje se podvrgavaju oksidaciji u tkivima viših životinja su ili ekstracelularna tečnost ili endogeni intracelularni lipidi. U mišićnom tkivu, uključujući srčani mišić, masne kiseline prolaze kroz direktnu oksidaciju. Glavni izvor endogenih masnih kiselina je rezervna mast sadržana u citoplazmi. Masne kiseline se prvo hidroliziraju specifičnim intracelularnim enzimima lipaze u glicerol i slobodne masne kiseline, a potonje se podvrgavaju aktivaciji i oksidaciji. Drugi izvor masnih kiselina su membranski fosfogliceridi. Metabolička obnova fosfoglicerida se stalno odvija, pri čemu nastaju slobodne masne kiseline.
Razgradnja masti.
Pljuvačka ne sadrži enzime za cijepanje masti. U želucu također nema primjetne probave masti iz hrane, ali dolazi do djelomične destrukcije lipoproteinskih kompleksa ćelijskih membrana hrane, što masti čini pristupačnijim za naknadno izlaganje lipazi soka pankreasa.
Razgradnja masti kod ljudi se događa uglavnom u gornjim dijelovima tankog crijeva, gdje postoje uslovi za emulgiranje masti. Najjače emulgirajuće dejstvo na masti imaju žučne soli koje sa žučom ulaze u duodenum u obliku soli natrijuma. Žučne kiseline su krajnji proizvod metabolizma holesterola. Po svojoj hemijskoj prirodi, žučne kiseline su derivati holanske kiseline. Žučne kiseline su prisutne u žuči u konjugiranom obliku. Ove veze se ponekad nazivaju uparene, jer. sastoje se od dvije komponente: žučne kiseline i glicina, te žučne kiseline i taurina. Žučne soli smanjuju površinsku napetost na površini vode i masti. Žučne kiseline takođe igraju važnu ulogu kao aktivator pankreasne lipaze, pod čijim uticajem dolazi do razgradnje masti u crevima. Lipaza koju proizvodi pankreas razgrađuje trigliceride koji su u emulgiranom stanju. Intestinalna lipaza je također uključena u razgradnju masti, katalizuje hidrolitičku razgradnju monoglicerida i ne djeluje na di- i trigliceride. Tako se u tankom crijevu glavni dio masti apsorbira nakon cijepanja na masne kiseline i glicerol, koji se, budući da su vrlo topljivi u vodi, apsorbiraju u crijevima, a zatim ulaze u krvotok portalne vene i odatle u jetra.
Dugolančane masne kiseline i monogliceridi se apsorbuju uz učešće žuči (žučne kiseline). Masne kiseline i monogliceridi formiraju micele koje su stabilne u vodenom mediju. Glavni endogeni izvor masnih kiselina je rezervna mast. Trigliceridi depoa masti igraju istu ulogu u metabolizmu lipida kao glikogen jetre u metabolizmu ugljikohidrata. Samo slobodne masne kiseline mogu se koristiti kao izvor energije, a trigliceridi moraju biti hidrolizirani lipazama u glicerol i slobodne masne kiseline. Mogu prijeći iz masnih depoa u krvnu plazmu i koristiti se kao energetski materijal.
Drugi izvor masnih kiselina su membranski fosfolipidi. Ćelije kontinuirano prolaze kroz metaboličku obnovu fosfolipida, pri čemu nastaju slobodne masne kiseline (fosfolipidi).
Oksidacija masnih kiselina u ćelijama odvija se u mitohondrijima uz učešće multienzimskog kompleksa . Proces oksidacije masnih kiselina sastoji se od sljedećih koraka:
Faza 1 - aktivacija masnih kiselina i njihov prodor iz citoplazme u mitohondrije, uz stvaranje acil-CoA. Uključuje 3 faze:
a) enzimska esterifikacija slobodne masne kiseline citoplazmatskim CoA (na račun ATP energije) u vanjskoj membrani mitohondrija.
b) transfer ostatka masne kiseline sa CoA na molekul karnitina, uz pomoć kojeg se ovaj ostatak transportuje kroz unutrašnju mitohondrijalnu membranu.
c) prijenos ostatka masne kiseline sa karnitina na intramitohondrijalni CoA.
Faza 2 - prva faza dehidrogenacije:. Kada se masne kiseline oksidiraju, to slijedi fazu aktivacije i događa se unutar mitohondrija. Acyl-CoA prolazi kroz enzimsku dehidrogenaciju, pretvarajući se u CoA ester nezasićene kiseline.
Faza 3 - faza hidratacije: dodavanje molekule vode i formiranje b-hidroksil CoA.
Faza 4 - druga faza dehidrogenacije, formira se b-ketoacil CoA.
Faza 5 - reakcija tiolaze: u njoj nastaje acetil CoA, koji se podvrgava oksidaciji u Krebsovom ciklusu, i acil CoA, koji zatim prolazi putem b oksidacije. Korak tiolitičkog cijepanja je tioliza, vrlo egzorganska reakcija.
Sekvencijalno oksidativno cijepanje acetil CoA molekula od CoA estera dugolančanih zasićenih masnih kiselina naziva se b oksidacija.
Energetski bilans: U slučaju oksidacije, na primjer palmitinske kiseline, G je 9797 kJ.
Najčešće kiseline povrća, bobičastog voća i voća su neisparljive limunske, jabučne, vinske i druge kiseline. Sadržaj kiselina u šumskom voću kreće se od 0,6 do 6%. Najveća kiselost je tipična za brusnicu, morsku krkavinu, viburnum.
Kiseline su uključene u formiranje okusa šumskog voća, smanjujući pH okoliša, povoljno utječu na proces probave, doprinose stvaranju određenog sastava mikroflore i inhibiraju procese propadanja u gastrointestinalnom traktu. Fenolne kiseline imaju baktericidni efekat.
Osim nehlapljivih kiselina, voće i bobice sadrže malu količinu hlapljivih kiselina: mravlje, valerijanske i kapronske. Svi su sadržani u sastavu viburnuma. Sorbinska i parasorbinska kiselina tipične su za plodove planinskog pepela. Mravlja kiselina se nalazi u malinama. Fenolne kiseline i njihovi estri nalaze se u malim količinama u različitim bobicama: na primjer, salicilna kiselina se nalazi u malinama, kupinama, jagodama, brusnicama, planinskom pepelu. Isparljive kiseline formiraju aromu svježeg bobičastog voća.
Lipidi. Sastav voća i bobičastog voća sadrži nešto lipida
(0,1-0,3%), koncentrisani su uglavnom u semenu. Izuzetak je morska krkavina, koja sadrži lipide i u pulpi i u sjemenkama. Koncentracija lipida u sjemenkama kreće se od 4 do 31%.
Lipidi igraju važnu ulogu jer su dio ćelijskih membrana. U voću i bobicama, lipidi su predstavljeni mastima, voskovima, kutinom, steroidima i drugim jedinjenjima. Masti voća i bobičastog voća karakteriše povoljan sastav masnih kiselina sa prevlašću nezasićenih masnih kiselina.
Voskovi, kutin, ursolna kiselina formiraju voštani premaz, karakterističan, na primjer, za borovnice i brusnice.
Vitamini. Nutritivna vrijednost povrća, voća i bobičastog voća u velikoj mjeri određuje prisustvo vitamina i supstanci sličnih vitaminima u njima. Voće i bobičasto voće sadrže vodu i vitamine rastvorljive u mastima. One rastvorljive u vodi uključuju askorbinsku kiselinu, vitamine B, bioflavonoide.
Vitamin C, tačnije njegov redukovani oblik - L-askorbinska kiselina je česta u samoniklom voću i bobičastom voću. Mnogi od njih, uz redovnu upotrebu, lako mogu popuniti dnevnu potrebu za askorbinskom kiselinom (50-100 mg dnevno).
Askorbinska kiselina je aktivan učesnik u mnogim redoks procesima koji se odvijaju u ljudskom tijelu. Poboljšava otpornost na zarazne bolesti, posebno prehlade, nepovoljni vanjski utjecaji (pregrijavanje, hlađenje), povećava efikasnost. Askorbinska kiselina utiče na hematopoezu, metabolizam ugljenih hidrata i holesterola. Biološka uloga vitamina C najpotpunije se očituje u prisustvu P-aktivnih spojeva, što je tipično za voće i bobičasto voće. Za osobu dnevna norma askorbinske kiseline je 70-100 mg, P-aktivnih tvari - 30-50 mg.
Bobice iz porodice brusnica, kao i kupine, bobice, ptičja trešnja sadrže 10-20 mg vitamina C / 100 grama. Njegova prosječna količina (od 30 do 75 mg / 100 grama) tipična je za jagode, viburnum, planinski jasen, borovnice.
Crna ribizla po sadržaju askorbinske kiseline nadmašuje mnoge vrste bobičastog voća, akumulirajući je do 500 mg / 100 grama.
Grupa P-aktivnih jedinjenja uključuje antocijanine, leukoantocijanine, katehine, flavonole, koji se razlikuju po hemijskom sastavu. P-aktivna jedinjenja imaju antihipertenzivne i antisklerotične efekte. Ukupan sadržaj P-aktivnih supstanci je 0,-0,6% u voću i bobičastom voću. U intenzivno obojenim kulturama i usevima aronije, kao što su borovnica, crna ribizla, borovnica, sadržaj P-aktivnih supstanci dostiže 1-1,5%.
B vitamini su uključeni u prostatičnu grupu enzima koji provode energetski metabolizam, biosintezu purinskih baza; regulišu metabolizam ugljikohidrata, proteina i masti. Od šumskog voća izvor riboflavina su šipak, borovnice, borovnice, a izvor tiamina je planinski pepeo.
Veliki i umjereni sadržaj riboflavina tipičan je za zeleni grašak, slatku papriku i karfiol.
Uz nedostatak tiamina i riboflavina, razvija se beriberi, koji se očituje povećanim umorom.
Predstavnici porodice vitamina B6 pronađeni su u mnogim šumskim plodovima.- piridoksin, piridoksal, piridoksamin, ali u malim količinama (od 0,1 do 3,9 mg / 100 grama). Fiziološka potreba odrasle osobe za ovim vitaminom je 1,8-2 mg dnevno. Njegov nedostatak dovodi do nervnih poremećaja, pospanosti, upale kože, očiju, dok se potreba za niacinom povećava, poremećena je upotreba aminokiselina i proteina.
Niacin (PP) je potreban osobi u značajnim količinama - 14-28 mg dnevno. Njegovim nedostatkom u ishrani javlja se razdražljivost, depresivno raspoloženje, glavobolja i drugi bolni simptomi. Niacin se nalazi u svim šumskim plodovima. Borovnice, šipak, morski trn i maline sadrže više niacina od ostalih.
Izvor vitamina grupe B i PP su i krompir, karfiol, pečurke - pečurke (28,15 mcg / 100 g) i lisičarke (25,38 mcg / 100 g).
Neke bobice su od najveće vrijednosti kao izvor folne kiseline. Ovaj vitamin je antianemični faktor, stimuliše stvaranje crvenih i belih krvnih zrnaca. Od bobičastog voća koje može ispuniti čovjekovu potrebu za ovim vitaminom, potrebno je izdvojiti planinski pepeo, divlju ružu, jagode, maline.
Od povrća, folna kiselina se nalazi u zelenom grašku, kupusu (20-40 mcg na 100 g proizvoda), a od gljiva - u belogu, pečurkama i šampinjonima (40 mcg; 30 mcg i 30 mcg, respektivno).
Vitamini rastvorljivi u mastima uključuju A, D, E, K. Šumsko voće sa intenzivnim žutim mesom bogato je karotenoidima, uključujući B-karoten, koji se u ljudskom organizmu najefikasnije pretvara u vitamin A. Glavni akumulatori B-karotena su morska krkavina, glog, šipak. Karotenoidi sprečavaju tkivnu hepaksiju, učestvuju u stvaranju hormona nadbubrežne žlezde, vizuelnog pigmenta.
Izvor ovih vitamina je povrće kao što su karfiol, šargarepa i paradajz.
Vitamin E stabilizira i štiti nezasićene lipide od pretjerane oksidacije, te sprječava neplodnost kod ljudi. Najveća biološka aktivnost je karakteristična za a-tokoferol. Tokoferol sintetiziraju samo biljke. Morska krkavina je izuzetno bogata njima - do 18 mg/100 grama u plodovima i deset puta više u ulju. Šipak, glog, planinski pepeo bogati su tokoferolima.
Vitamin K podstiče zgrušavanje krvi. U bobicama se nalazi u obliku K1. Više od ostalih bobica, vitamin K1 akumulira planinski pepeo, crna ribizla, divlja ruža, borovnica.
LIPIDI
Lipidi (od grčkog - masno) su prirodni nepolarni spojevi koji su potpuno ili gotovo potpuno netopivi u vodi, ali topljivi u nepolarnim otapalima, na primjer, kloroform, ugljični disulfid, eter i vrući etanol. Glavna karakteristika ove grupe prilično heterogenih supstanci je njihova nepolarnost, što se u nekim slučajevima objašnjava prisutnošću u ovim spojevima jedne ili više masnih kiselina koje sadrže duge alifatske ugljikovodične lance. Sadržaj lipida u tijelu je 10 - 20% tjelesne težine.
Lipidi obavljaju sljedeće biološke funkcije :
1. One su komponente membrana koje u velikoj mjeri određuju njihovu propusnost.
2. Služe kao jedan od glavnih oblika skladištenja ugljenika i energije (1 g masti tokom oksidacije daje 9,3 kcal).
3. Mogu biti prekursori drugih važnih jedinjenja.
4.Obavljaju ulogu zaštitnih barijera koje štite od toplotnih, električnih i fizičkih uticaja.
5. Oni su dio zaštitnih ljuski koje štite od infekcije i prekomjernog gubitka ili nakupljanja vode. Masti su izvori endogene vode.
6. U nekim slučajevima to su vitamini i hormoni.
7. Učestvuju u smanjenju nervnih impulsa, jer su deo mijelinskih ovojnica.
Postoji klasifikacija zasnovana na sadržaju jednog ili drugog alkohola u sastavu masti. Shodno tome, izolovani su lipidi i holesterol koji sadrže glicerin, sfingozin i njegovi derivati.
Postoji klasifikacija lipida prema njihovoj strukturi:
1. Jednostavni lipidi. 2. Kompleksni lipidi. 3. Lipidni derivati.
Jednostavne masti uključuju samo estre masnih kiselina i alkohola. Sastav složenih lipida, pored masnih kiselina i alkohola, uključuje i druge komponente. Među složenim lipidima razlikuju se fosfoacilgliceroli, sfingomijelini, cerebrozidi i gangliozidi. Derivati lipida uključuju sva jedinjenja koja se ne mogu jasno klasifikovati kao jednostavne ili složene lipide, kao što su steroidi, karotenoidi i vitamini lipidne prirode. Mnogi od njih su formirani od ugljikovodičnih lanaca koji su bili dio masnih kiselina.Sastav jednostavnih i složenih lipida obavezno uključuje masne kiseline.U stvari, to su karboksilne kiseline sa dugim alifatskim lancem. Prirodne masne kiseline su veoma raznovrsne.
Radi praktičnosti, razlikuju se sljedeće grupe: 1. Većina masnih kiselina su monokarboksilne kiseline koje sadrže linearne ugljikovodične lance s parnim brojem atoma (obično C 12 - C 20). Manje uobičajene su FA sa kraćim lancima ili s neparnim brojem atoma ugljika. 2. Često postoje kiseline koje sadrže etilenske (nezasićene, dvostruke) veze (obično C 18 - C 20 kiseline). Ako postoje dvije ili više dvostrukih veza, one su gotovo uvijek odvojene jednom metilenskom grupom: - CH = CH - CH 2 - CH = CH -.
3. U nezasićenim kiselinama sa dvostrukim vezama, gotovo uvijek se odvija cis konfiguracija.
Veliki broj nepolarnih C - C i C - H veza u ugljičnom lancu daje suštinski nepolarni karakter cijeloj molekuli u cjelini, iako ima i polarnu grupu - COOH. Sasvim je jasno da će spoj koji sadrži jedan ili više FA ostataka biti nepolaran, što je razlog netopivosti lipida u vodi, a također uzrokuje sklapanje lipida u biomembrane.
Glavne prirodne masne kiseline koje čine tkiva:
Zasićeni LCD-i:
1. Laurić (C 12). 2. Miristic (C 14). 3. Palmitić (C 16).
4. Stearić (C 18). 5. Arachinoic (C 20).
II. Zasićeni LCD-i:
1. Palmitoleic (S 16:1) ∆9. 2.Oleić (S 18:1) ∆9. 3. Linolna (S 18:2) ∆9, 12
4. Linolenska (S 18:3) ∆9, 12, 15. 5. Arahidonska (S 20:4) ∆5, 8, 11, 14
Gdje je (S x:y) ∆ Z- x je broj atoma ugljika. y je broj dvostrukih veza.
Z je položaj dvostrukih veza (broj dolazi od - COOH).
FA se mogu oksidirati enzimski i neenzimski.
Neenzimska oksidacija masnih kiselina. (LPO) - na dvostrukim vezama oksidiraju se u hidroperoksi kiseline pod djelovanjem jakih oksidacijskih sredstava, na primjer, H 2 O 2; Oko 2; HE. Nastala jedinjenja peroksida su toksični oblici jedinjenja kiseonika. Složeni lipidi prisutni u biološkim membranama vrlo lako formiraju perokside, jer sadrže mnogo nezasićenih masnih kiselina. To, pak, može dovesti do oksidacije membranskih proteina, što može imati dubok utjecaj na strukturu i funkciju membrane. Takav destruktivni efekat u prvoj fazi izaziva pretvaranje lipidnih peroksida u peroksid - radikale - visoko reaktivne agense.Ova reakcija se može odvijati uz učešće iona metala, na primer gvožđa. Užeglost i kvarenje prehrambenih proizvoda dijelom su posljedica ovih reakcija.Žive ćelije su u stanju da se brane od djelovanja ovih oksidansa. Neutralizacija se odvija uz učešće glutationa, vitamina E i C koji djeluju kao antioksidansi Enzimska oksidacija masnih kiselina - to je oksidacija masnih kiselina sa dugim lancem, posebno arahidonske kiseline, prekursora sinteze prostaglandina. Uopšteno, spojevi formirani od C 20 - polinezasićenih kiselina nazivaju se eikozanoidi. Arahidonska kiselina nastaje od membranskih fosfolipida, koji se pod dejstvom fosfolipaze A 2 razlažu na lizofosfolipide i arahidonsku kiselinu. Linolna kiselina takođe može biti prekursor arahidonske kiseline. Prostaglidini (PG) su dobili svoje ime jer su prvi put izolovani iz sjemene plazme proizvedene u prostatnoj žlijezdi. Sada je poznato da su prisutni u većini tkiva sisara, međutim, u izuzetno malim količinama (≤10 -9 g).
Prostaglandini regulišu sledeće procese u ćelijama :
1. Redukcija glatke mišiće.
2. Cirkulacija krvi (krvni pritisak).
3. Prijenos nervnih impulsa.
4. Učestvujte u zapaljenom procesu.
5. Ravnoteža vode i elektrolita.
6. Zgrušavanje krvi.
Mehanizam djelovanja prostaglandina povezan je s prisustvom u ciljnim stanicama koje kontroliraju prostaglandini specifičnih membranskih proteina – receptora koji se vezuju za određene prostaglandine. Konačni efekat je posredovan djelovanjem cikličkih nukleotida – prostaglandina povećava ili smanjuje (u zavisnosti od prirode tkiva) sadržaj intracelularnog cAMP ili cGMP. Postoji 6 glavnih tipova PG. Generale strukturnu formulu prostaglandina pokazuje da je to C 20 monokarbonska kiselina sa ciklopentanskim prstenom unutar lanca. Različiti prostaglandini imaju jednu ili dvije dvostruke veze na određenim pozicijama i također se vezuju na određenim pozicijama za atome kisika.
Biosinteza prostaglandina počinje pretvaranjem arahidonske kiseline u dva endoperoksidna intermedijera - PGG 2 (prvi formiran) PGH 2 . Obje reakcije katalizira isti enzim - prostaglandin endoperoksid sintaza, koja se sastoji od dvije komponente - ciklooksigenaze (adicija O 2 i formiranje ciklopentanskog prstena) i peroksidaze (odgovorne za formiranje hidroperoksidnih grupa). U nekim ćelijama, PGG 2 i PGH 2 se pretvaraju u druge proizvode, tromboksane i prostociklin. Formiranje tromboksana se javlja u trombocitima i potiče zgrušavanje krvi, uzrokujući agregaciju trombocita i sužavanje arterija. Naprotiv, stvaranje prostociklina događa se u stanicama koje oblažu arterije i vene i sprječava zgrušavanje krvi inhibirajući iste procese. Neravnoteža ovih jedinjenja u korist agregacije trombocita može biti osnovni uzrok stvaranja adsorbentnih plakova. Biohemija prostaglandina objašnjava terapeutsko dejstvo aspirina, koji služi kao efikasan lek za ublažavanje blagih bolova, upala, kao antipiretik, tj. ona stanja, čija pojava u određenoj meri zavisi od povećane sinteze prostaglandina. Aspirin smanjuje brzinu njihove sinteze, djelujući kao inhibitor ciklooksigenazne komponente sinteze endoperoksida, dok dolazi do kovalentne modifikacije važne bočne amino grupe enzima. Drugi protuupalni agens, indometozin, također inhibira sintezu, ali ne kovalentnom modifikacijom. Lijekovi steroidne prirode djeluju na drugačiji način, na primjer, inhibirajući stvaranje arahidonske kiseline iz membranskih lipida. Nedavno je otkrivena nova klasa jedinjenja, nazvana leukotrieni. Sintetiziraju ih leukociti i sadrže tri konjugirane dvostruke veze. Ovo su snažni aktivatori kontrakcije glatkih mišića. Na njih su posebno osjetljive male bronhiole, što je vjerovatno posljedica otežanog disanja kod pacijenata bronhijalna astma. Reumatoidni artritis je takođe posledica upalnog dejstva leukotriena. Sinteza leukotriena također počinje arahidonskom kiselinom, ali u ovom slučaju na nju utiče enzim lipoksigenaza, koji pretvara arahidonsku kiselinu u hidroperoksid. Ni aspirin ni indometocin ne utiču na ovaj enzim. Najmoćniji leukotrieni su jedinjenja sa tioeterskom vezom: - C - S - C - nastala kao rezultat kondenzacije sa glutationom.
Razmotrite različite vrste lipida:
I. Jednostavni lipidi .
Jednostavni lipidi se dijele u dvije grupe:
1. Neutralni acilgliceroli.
1. Neutralni acilgliceroli.
U zavisnosti od broja FA ostataka vezanih za trihidrični alkohol glicerol, razlikuju se mono-, di-, triacil-gliceroli. Triacil derivati su najčešći u prirodi. U svim slučajevima, prosti acilglicerol ne sadrži funkcionalne jonske grupe i pripada neutralnim lipidima. Acil bočni lanci su obično različiti.
Opća formula:
CH 2 O - C - R
CHO - C - R /
CH 2 O - C - R //
TAG-ovi obavljaju tri glavne funkcije u životinjskim tkivima:
1. U masnom tkivu se talože u rezervi (kao oblik skladištenja energije i ugljenika). Ovo su glavni lipidi rezervnih masti, ima ih i do 99%.
2. u obliku lipoproteinskih čestica (na primjer, hilomikroni) se transportuju duž limfni sistem i krvotok, koji se distribuira u sva tkiva (u ovom slučaju se transportuju oni TAG-ovi koji su sintetizirani iz probavljenih masnih kiselina).
3. TAG-ovi djeluju kao fizička zaštita i regulator temperature za različite organe tijela. Najčešće (50%) sadrže oleinsku kiselinu, 23% - palmitinsku, 10% - linolnu, 6% - stearinsku, 5% - palmitoleinsku i 6% - ostatak masnih kiselina.
TAG-ove karakteriziraju masne kiseline s parnim brojem atoma ugljika, ali postoje i masne kiseline sa neparnim brojem atoma ugljika - njih je 2-5%, a masnoće kože imaju 15-20%. Zasićenost određuje tačku topljenja masti. Potkožna mast je zasićenija od masti jetre. Sastav masnih kiselina lipida može varirati od T 0 sredine.
2.Voskovi.
To su estri, međutim i alkohol (do C18) i kiselina uključena u njihov sastav imaju duge ugljikovodične lance. Svi voskovi su potpuno nerastvorljivi u vodi. Prirodni voskovi su obično krajnji produkti onih metaboličkih puteva čija je glavna uloga stvaranje zaštitnih premaza. Ptičje perje i životinjska koža imaju voštani premaz koji ih čini vodoodbojnim. Voštani premaz listova i plodova biljaka smanjuje gubitak vlage i smanjuje mogućnost infekcije.
II Kompleksni lipidi.
Postoje tri glavne klase složenih lipida:
1. Fosfoacilgliceroli. 2. Sfingomijelini. 3. Glikolipidi.
Prve dvije klase nazivaju se fosfatidi ili fosfolipidi zbog prisustva fosfatnih grupa. Sva ova jedinjenja nalaze se samo u membranama, jer su njihove glavne komponente.
1. Fosfoacilgliceroli (FAG)
Ovo su najčešći složeni lipidi. Broj različitih FA i njihova relativna koncentracija uvelike varira u zavisnosti od vrste ćelija i njihovog fiziološkog stanja. Uprkos ovoj raznolikosti, oni i dalje razlikuju glavne i sporedne komponente. Glavni: fosfatilholin (lecitin), fosfatidiletanolamin (cefalin), fosfatidilglicerin. manji: fosfatidilserin, difosfatidilglicerol (kardiolipid), fosfatidilinozitol i fosfatidna kiselina. Svi ostali FAG se formiraju od fosfatidne kiseline.
CH 2 O - C - R
CHO - C - R 1
CH 2 O - R - R 1
Fosfatna kiselina.
Različiti FAG-ovi se međusobno razlikuju po dodatnim grupama vezanim fosfoesterskom vezom za fosfatidnu kiselinu. Sastav FA različitih FAG-ova se razlikuje čak i unutar istog organizma. Neki lecitini, kao što je dipalmitoilecitin, glavni su element surfaktanta (surfaktanta) pluća – njegov nedostatak kod prijevremeno rođenih beba uzrokuje respiratornu insuficijenciju.
Struktura nekih FA:
CH 2 O - C - R CH 2 O - C - R
CHO - C - R1 CHO - C - R 1
CH 2 O - R - OH CH 2 O - R - OH
O - CH 2 - CH 2 - N + H 3 O - CH 2 - CH 2 - N + - (CH 3) 3
Fosfatidiletanolamin Fosfatidilholin
(cefalin) (lecitin)
CH 2 O - C - R CH 2 O - C - R
CHO - C - R 1 CHO - C - R 1
CH 2 O - R - OH N + H 3 CH 2 O - R - OH OH
O - CH 2 - CH - COOH O - CH 2 - CH - CH 2 OH
Fosfatidilserin Fosfatidilglicerin
CH 2 O - C - R O CH 2 O - C - R
| || R /// - C - OH 2 C ||
CHO - C - R 1 O | CHO - C - R 1
| || R // - C - ONS ||
CH 2 O - R - OH O | CH 2 O - R - OH
O - CH 2 - CH - CH 2 O - R - OH 2 C O
|
|
Difosfatidilglicerol Fosfatidilinozitol
(kardiolipid)
On čini do 15% svih lipida miokarda.Ponekad se u fosfoacilglicerolima jedna od masnih kiselina zamjenjuje aldehidom, pri čemu nastaju plazmalogeni.
CH 2 - O - CH \u003d CH - R /
CH - O - C - R 2
CH 2 - O - R - OH
U ovim strukturama se jasno vidi da jedinjenja imaju dva različita regiona: nepolarni hidrofobni „rep“ i polarnu hidrofilnu „glavu“. Takva dvojna priroda naziva se alifatska (od grčkog - dvostrano i osjećanje).
2.Sfingomijelini (fosfosvingolipidi).
Sastoje se od jedne molekule masnih kiselina, holina, fosforne kiseline i swingozin alkohola. Sfingozini su porodica dugih nezasićenih lanaca amino alkohola koji se razlikuju po dužini lanca. FA je vezan za amino grupu sfingozina preko amidne veze i nastaje ceramid. Dodavanje ostatka fosforilholina dovršava formiranje swingomijelina:
CH 3 - (CH 2) 12 - CH \u003d CH - CH - CH - CH 2 O - R - O - CH 2 - CH 2 - N + (CH 3) 3
Sfingomijelini
Sfingomijelini takođe imaju amfipatska svojstva. Sfingomijelini se nalaze u značajnim količinama u ćelijskim membranama nervnog i moždanog tkiva.
3. Glikolipidi (glikosfingolipidi).
Takođe su na bazi ceramida. Postoje cerebrozidi i gangliozidi. Još jedna sekvencijalna reakcija biosinteze lipida je enzimska konverzija prekursora ceramida u cerebrozide i gangliozide vezivanjem grupe ugljikohidrata na ceramid. Cerebrozidi i gangliozidi se uglavnom (iako ne isključivo) nalaze u ćelijskim membranama nervnog i moždanog tkiva i takođe pokazuju amfipatska svojstva. Protozoa cerebrozidi sadrže monosaharidnu grupu vezanu glikozidnom vezom za krajnju -OH grupu ceramida. U pravilu je to glukoza ili galaktoza.
CH \u003d CH - (CH 2) 12 - CH 3
Oko 25% cerebrozida može biti predstavljeno sulfoniranim (na 3. poziciji) galaktocerebrozidima. Drugi cerebrodidi sadrže duže oligosaharidne grupe (2-10 ostataka) koje sadrže amino šećere, N-acilsaharide i fruktozu. Gangliozidi također sadrže oligosaharidnu grupu, ali ih karakterizira prisustvo najmanje jednog ostatka sijalinske kiseline. Jedan gangliozid se razlikuje od drugog (ovo se odnosi i na cerebrozide) po strukturi oligosaharidne komponente. Ostaci sijalične kiseline u gangliozidima sadrže od 1 do 4.
Funkcije glikolipida:
1. Davanje krutosti membranama.
2. Formiranje antigenskih hemijskih markera ćelije, određivanje krvne grupe (prema ABO sistemu).
3. Formiranje hemijskih markera faza diferencijacije ćelija.
4. Regulacija normalnog rasta ćelija i samim tim je povezana sa transformacijom normalnih ćelija u tumorske ćelije.
5. Davanje ćelijama sposobnosti da reaguju sa nekim biološki aktivnim jedinjenjima, na primer, sa bakterijskim toksinima (kolera, tetanus), glikoproteinskim hormonima, interferonima i virusima, tj. u stvari, to je funkcija receptora. Svi ovi procesi su od velikog biološkog značaja. Uz glikolipide, glikoproteini također učestvuju u mnogim od ovih procesa. Ovo naglašava važnost oligosaharida za procese koji se odvijaju na površini membrane.
6.učestvuju u međućelijskim interakcijama.
III. Lipidni derivati.
Ovo je veoma heterogena grupa jedinjenja. Sličnost je ograničena samo slabom rastvorljivošću u vodi. Najvažniji članovi ove grupe su steroidi, karotenoidi i srodni vitamini lipidne prirode.
1. Steroidi. Nalaze se u svim organizmima gdje obavljaju različite funkcije. Kod ljudi igraju ulogu polnih hormona, emulgatora u varenju lipida i uključeni su u transport lipida kroz membrane. U krvnoj plazmi djeluju kao protuupalni agensi i kao regulatori nekih metaboličkih procesa. Svi steroidi imaju sličnu strukturu, koja se zasniva na perhidrociklopentanofenantrenu. Raznovrsnost steroidnih struktura je posledica različitog stepena nezasićenosti i prisustva nekoliko grupa u različitim pozicijama ciklusa.
 |
Perhidrociklopentanofenantren
Prisustvo bočnog lanca ugljikovodika (C 8 - C 10) na 17. poziciji i hidroksilne grupe na poziciji 3 karakteristično je za veliku grupu steroida zvanih steroli. Najvažniji i najčešći od njih je holesterol. Ulazi kao strukturna komponenta u ćelijske membrane, međutim njegova koncentracija u membranama varira (0 - 40% ukupnog sadržaja lipida u membranama). Kolesterol (zbog sadržaja konsolidirane strukture koja je manje fleksibilna od produženog lanca ugljikovodika) daje membrani veću krutost (snagu). Kolesterol je također primarni metabolički prekursor drugih važnih steroida, uključujući žučne kiseline i polne hormone. U nekim tkivima holesterol je prekursor vitamina D. Spolni steroidni hormoni: muški - testosteron (uzrokuje sve polne karakteristike) i masni: estradiol i progesteron (uglavnom odgovorni za regulaciju menstrualnog ciklusa).
CH - (CH 2) 3 - CH - (CH3) 2
Holesterol
Prvo je izoliran iz žučnih kamenaca, pa otuda i naziv (chole - žuč, steros - tvrd).
2. Karotenoidi. Karotenoidi se dijele u dvije grupe: karoteni i ksantofili. Obje grupe uključuju pigmente netopive u vodi koji su široko rasprostranjeni u prirodi, a većina ih se nalazi u biljkama i algama. Karoteni su čisto ugljikovodična jedinjenja, dok ksantofili sadrže kisikove grupe. Karoteni su šire rasprostranjeni u prirodi. Najčešći je beta-karoten. To je C 40 ugljovodonik sa jako razgranatom strukturom i nezasićenim vezama, na oba kraja ugljovodoničnog lanca nalaze se identični supstituisani prstenovi. Svi ostali karotenoidi, u suštini, mogu se smatrati varijantama ove strukture. Prilikom enzimatskog cijepanja beta-karotena simetrično nastaju dvije molekule vit A. U mrežnjači oka redukovani alkoholni oblik vitamina A (retinol) enzim pretvara u oksidirani aldehidni oblik (retinal), koji stvara kompleks sa različitim proteinima koji se vezuju za retinu (opsini). Takav kompleks je primarni svetlosni receptor u fotosenzitivnim ćelijama, koje zatim prenose informacije do ćelija nervnog sistema. Postoje dvije vrste fotosenzitivnih ćelija:
1. Štapići - percipiraju slabo svjetlo i neosjetljivi su na boje.
2. Češeri - percipiraju jako svjetlo i odgovorni su za formiranje slike u boji.
Ispostavilo se da u štapićima postoji samo jedan oksin (rodopsin). Češeri sadrže najmanje 3 vrste toksina. Oni formiraju komplekse sa retinalom kako bi formirali receptore osjetljive na plavu, crvenu i zelenu boju. Kombinacija ovih boja daje različite nijanse. Ukupno, ljudsko oko može razlikovati 256.000 nijansi. Utvrđeno je da se retinal kovalentno vezuje za opsin, formirajući kompleks rodopsin-retina. U ovom slučaju je vezan cis-oblik retine, koji kroz mnoge međufaze prelazi u trans-izomer. U ovom slučaju, jedan ili više međuformi tada učestvuju u stvaranju vrlo malih električnih potencijala koji se aktiviraju nervni sistem. Ciklus transformacija fotosenzitivnog pigmenta tokom vizuelne percepcije završava se regeneracijom cis-retinala.
Lipidni vitamini.
3.Vitamin D. Potreban za normalan metabolizam kalcijuma i fosfora neophodnih za rast zdravih kostiju i zuba. Njegov nedostatak uzrokuje rahitis - bolest u kojoj kosti postaju meke i plastične, što dovodi do njihove deformacije. Vitamin D nastaje iz prekursora sterola uz UV zračenje. Jedan od važnih prekursora je 7-dihidroholesterol, koji se i sam dobija enzimskim putem iz holesterola. Iz njega se dobija vitamin D3. Postoji nekoliko oblika vit D. Fora D 3 (kolikalceferol) je prisutan u mlijeku, puteru, ribljoj jetri, koji su glavni egzogeni izvor ovog vitamina. Vitamin D 3 može se sintetizirati u koži, pod uslovom da dobije normalan dio sunčeve svjetlosti. I endogeni i egzogeni vit D 3 se transportuje plazmom do jetre i bubrega, gdje se pretvara u još aktivnije hidroksilirane derivate. Glavni aktivni oblik u tijelu je derivat 1,25-dihidroksi.
4.Vitamin E. Osnovna struktura vitamina E (a postoji nekoliko oblika) naziva se tokoferol.Najaktivniji oblik vitamina E je alfa-tokoferol. Kod pacova, vit E je neophodan za reprodukciju (ovo još nije dokazano za ljude). Kod nekih životinja nedostatak vit E dovodi do mišićne distrofije. Vit E se obično dodaje tokom prerade hrane, jer ima antioksidativna svojstva, sprečava kvarenje hrane usled oksidacije. U živim ćelijama, vit E također djeluje kao antioksidans, zajedno s vitaminom C i glutationom.
5. Vitamin K. Njegov nedostatak usporava zgrušavanje krvi. Njegova struktura je bazirana na biciklici naftohinon sistem za koji je vezan dugačak ugljikovodični lanac. Može doći iz hrane (svježe povrće, sir), ili ga mogu stimulirati bakterije u gastrointestinalnom traktu. Njegov koagulacijski učinak je određen učešćem u sintezi protrombina, koji potom prelazi u trombin.
Biomembrane.
Biomembrane zatvaraju ćeliju i mnoge organele unutar ćelije. To su vrlo aktivni biološki sistemi odgovorni za procese kao što su selektivni transport supstanci u i iz ćelije i subcelularnih odjeljaka (formacija), vezivanje hormona i drugih regulatornih molekula, reakcije katalizirane enzimima, prijenos električnih impulsa i čak i sintezu ATP-a. Membrane se razlikuju jedna od druge, a različite vrste membrana imaju različite vrste funkcionalne aktivnosti. Na primjer, samo je unutrašnja mitohondrijska membrana odgovorna za biosintezu ATP-a.
Hemijski sastav: Membrane se sastoje od molekula lipida i proteina. Njihova relativna količina se značajno razlikuje za različite membrane, varirajući od 20% proteina + 80% lipida do 75% proteina + 25% lipida. Ugljikohidrati u obliku glikoproteina i glikolipida čine 0,5-10% membranske tvari.
lipidni dvosloj. Biomembranski lipidi su raspoređeni u dva sloja. Svaki monosloj se sastoji od složenih lipida (a ponekad i holesterola) raspoređenih na takav način da su nepolarni hidrofobni repovi molekula u bliskom kontaktu jedni s drugima; u istom kontaktu - polarni hidrofilni elementi. Sve ove interakcije su isključivo nekovalentne. Kombinuju se dva monosloja, orijentisani „rep“ prema „repu“, tako da se formira dvoslojna struktura koja ima nepolarni unutrašnji deo i dve polarne površine. Debljina svakog dvosloja je ≈ 3,5 - 4,0 nm. Sastav lipida monoslojeva je različit. Na primjer, cerebrozidi i gangliozidi se obično nalaze u vanjskom monosloju ćelijske membrane. Stepen ove razlike u lipidnom sastavu monoslojeva je različit za različite membrane i može se mijenjati kako stanica učestvuje u različitim procesima i kako stari. Molekuli lipida mogu se kretati iz jednog monosloja u drugi. Pokretljivost (rigidnost) i fluidnost lipidnog dvosloja određena je vrstom i dužinom ugljikovodičnih grupa koje čine masne kiseline i sfingozine, kao i sadržajem kolesterola. Povećana krutost je određena povećanim omjerom zasićenih i nezasićenih lanaca i povećanim sadržajem kolesterola. Fizička svojstva membrana također zavise od vrste i rasporeda proteina povezanih s lipidnim dvoslojem.
Membranski proteini . Ukupna struktura membrane uključuje proteine vezane za lipidni dvosloj na jedan od dva načina:
1.povezan sa hidrofilnom površinom dvosloja - (proteini površinske membrane);
2. uronjeni u hidrofobnu zonu lipidnog dvosloja - (integralni membranski proteini).
Svaka membrana sadrži oba tipa proteina, međutim, različite vrste membrana imaju različite omjere ovih proteina. Površinski proteini sa svojim polarnim bočnim grupama aminokiselinskih ostataka koji se nalaze na površini proteinske molekule su nekovalentno vezani za hidrofilnu površinu lipidnog dvosloja. Integralni proteini se razlikuju po stepenu uranjanja u hidrofobnu regiju dvosloja. Na obje strane membrane neki proteini samo ne mogu prodrijeti duboko u dvostruki sloj, dostižući otprilike polovicu sloja, dok drugi proteini svojom strukturom, takoreći, probijaju dvostruki sloj. Proteini, poput lipida, mogu promijeniti svoju lokaciju u membrani. Ovo kretanje može biti usmjereno duž monosloja, može biti rotacijsko unutar monosloja ili čak dvosloja, može biti usmjereno s jedne površine dvosloja na drugu. Proteini mogu biti podvrgnuti promjenama u svojoj konformaciji, što može utjecati na konformaciju susjednih proteina.
Funkcije membranskih proteina : Svi membranski proteini (i površinski i integralni) mogu se podijeliti u 2 velike grupe na osnovu njihove uloge u sastavu membrana:
1. strukturni proteini;
2. dinamički proteini.
Strukturni proteini pomažu u održavanju strukture cijele membrane. Često proteini ovog tipa imaju izdužen oblik i nalaze se na hidrofilnoj površini lipidnog sloja, djelujući kao molekularna traka. Dinamički proteini su proteini koji su direktno uključeni u ćelijske procese koji se odvijaju na membrani. U svim tipovima ćelija obično se razlikuju tri klase takvih proteina:
1. transportni proteini (učestvuju u transportu jedinjenja u i iz ćelija;
2. katalitički proteini (igraju ulogu enzima u reakcijama koje se odvijaju na membrani);
3. proteini - receptori (specifično vezuju određena jedinjenja - hormone, toksine, neurotransmitere na spoljnoj strani membrane, što služi kao signal za promenu hemijskih procesa u membrani ili unutar ćelije).
membranski transport.
Prenos (transport) jedinjenja unutar ili van ćelije, kao i njihov transport između citoplazme i raznih subcelularnih organela (mitohondrija, jezgra, itd.) određen je membranama. Transportna svojstva membrane određuju njenu polupropusnost - neki spojevi mogu prodrijeti kroz nju, dok drugi ne. Ovisno o specifičnom spoju i vrsti membrane, odvija se pasivni ili aktivni transport.
1 .Pasivni transport je transport duž gradijenta koncentracije bez energetske potrošnje ćelije. Takav transport se ponekad naziva difuzijom. Razlikovati jednostavnu i olakšanu difuziju.
Jednostavna difuzija se dešava bez ikakve interakcije sa membranskim proteinom. Prenos supstanci kroz lipidne dvosloje vrši se kroz pore u membrani formirane prazninama u dvosloju, ili kroz kanale formirane od membranskih proteina. Tako se, na primjer, prenose voda, neki organski joni i neka jedinjenja lipidnog tipa. Olakšana difuzija uključuje molekule nosače, obično membranski protein. Takav proces obično počinje vezivanjem prenesenog jedinjenja za transportni protein. Transportni protein se približava suprotnoj strani membrane, oslobađa ovo jedinjenje, preuzima prvobitno stanje i ponovo je spreman za obavljanje svojih funkcija. Druga verzija prijenosa: protein nije u stanju proći kroz dvosloj, a vezano jedinjenje prelazi s jednog proteina na drugi dok se ne nađe na suprotnoj strani membrane i tamo se oslobodi.
2 Aktivni transport je slučaj kada prijenos zahtijeva utrošak energije ćelije. Također se koristi ATP energija i drugi izvori energije. U ovom slučaju neophodno je i učešće membranskih proteina, a transportni sistem se može pokazati multimolekularnim. Energija se može potrošiti u bilo kojoj fazi prijenosa. U nekim slučajevima, jedan od stadija može biti stimulisan hormonom.
Jedan od najvažnijih sistema - K+, Na+ - pumpa. Pumpa je jedan protein koji se zove Na +, K + - ATPaza. Molekul ovog proteina prožima cijelu membranu i predstavlja oligomer s molekulskom težinom od 250.000. Mehanizam djelovanja uključuje nekoliko faza:
1. ATP molekul iz citoplazme vezuje se za aktivno mjesto na jednoj od podjedinica ATPaze. Vezivanje ATP-a je praćeno vezivanjem 3 Na+ iz citoplazme.
2. Fosforacija ATPaze (zbog ATP-a) uzrokuje konformacionu tranziciju u strukturi proteina, što dovodi do njegove preorijentacije na način da se vezani Na+ izlučuje kroz kanal otvoren prema vanjskoj strani membrane.
3. Nakon uklanjanja Na+, 2K+ izvana se vezuju za centre za vezivanje jona fosforizovanog proteina.
4. Uklanjanje fosfatne grupe hidrolizom uzrokuje novu konformacijsku tranziciju, vraćajući prvobitnu konformaciju, koja oslobađa 2K+ u kanal, sada otvoren unutar ćelije.
Dakle, za svaki potrošeni ATP molekul, ATPaza prenosi 3Na+ i 2K+ (odnosno iz ćelije i u ćeliju).
Mišićne ćelije sadrže Ca ++ - ATPaznu pumpu koja kontroliše kontrakciju mišića, a oslobađanje Ca ++ izaziva kontrakciju mišića.
membranskih receptora. Odgovor živih ćelija na izlaganje razne vrste biološki aktivnih supstanci (hormoni, razni toksini, neurotransmiteri itd.) počinje vezivanjem tvari za vanjsku površinu ćelijske membrane na određenom mjestu koje se zove receptor. Obično su to proteini, ali mogu biti i druge supstance, kao što su gangliozidi. Razmotrite rad nekih receptora.