Fiziološke funkcije ljudskog tijela. Fizička i fiziološka svojstva skeletnih, srčanih i glatkih mišića. Udžbenik "opšta fiziologija"
Predavanje 1
Uvod u kurs
Predmet i zadaci fiziologije
namjerno stvaraju nove lijekove. Stoga studenti koji studiraju farmaceutsku hemiju moraju poznavati savremene ideje o mehanizmima farmakološkog i toksičnog djelovanja na organizam i biti sposobni primijeniti stečeno znanje u budućim proizvodnim aktivnostima.
Osnovni fiziološki koncepti
Svaki naučna disciplina ima svoj konceptualni aparat. Naravno, fiziologija, nauka koja je nastala još u 19. veku, ima svoje koncepte i termine. Ispod su one najčešće. Ostali pojmovi i pojmovi koji imaju uže značenje biće razmatrani u toku izlaganja relevantnih dijelova fiziologije u narednim predavanjima.
Osnovni fiziološki koncepti.
Funkcija- specifična aktivnost sistema ili organa. Na primjer, funkcije gastrointestinalnog trakta su motorna, sekretorna, apsorpciona; funkcija respiratornog sistema je izmjena CO 2 i kiseonika; funkcija cirkulacijskog sistema je kretanje krvi kroz žile; funkcija miokarda - kontrakcija i opuštanje itd.
Proces- skup uzastopnih radnji ili stanja usmjerenih na postizanje određenog rezultata.
Mehanizam- način na koji se kontroliše proces ili funkcija. ‚ fiziologija se obično razlikuje nervozan i humoralni(tj. oslobađanjem hormona ili drugih bioloških kemikalija aktivne supstance) mehanizmi regulacije.
R regulacija- promjena intenziteta i smjera funkcije (procesa) kako bi se osigurala optimalna aktivnost organa i sistema.
Sistem u fiziologiji, to je skup organa, tkiva ili drugih strukturnih formacija povezanih zajedničkom funkcijom. Na primjer, kardiovaskularni sistem obezbjeđuje, uz pomoć srca i krvnih sudova, isporuku hranljivih, regulatornih, zaštitnih supstanci i kiseonika u tkiva, kao i uklanjanje metaboličkih produkata; izvozni (sekretorni) sistem ćelije obezbeđuje, uz pomoć endoplazmatskog retikuluma i Golgijevog aparata, sintezu i transport hormona, lipoproteina i drugih tajni iz ćelija.
OD tajna- specifičan proizvod vitalne aktivnosti ćelije koji obavlja određenu funkciju i oslobađa se u unutrašnje okruženje organizma. Proces stvaranja i izolacije tajne se zove sekrecija. Po sastavu se tajna dijeli na proteinsku (seroznu), sluzavu (mukoidnu), mješovitu i lipidnu.
Stimulus- faktori spoljašnje i unutrašnje sredine ili njihova promena, koji utiču na organe i tkiva, što dovodi do promene aktivnosti ovih poslednjih. Postoje mehanički, električni, hemijski, temperaturni, zvučni itd. podražaji. Stimulus može biti prag, tj. imaju minimalan efektivan uticaj; maksimum, čije daljnje povećanje snage ne dovodi do povećanja odgovarajuće reakcije; nadjačana, čije djelovanje može imati štetno ili bolno djelovanje ili dovesti do neadekvatnih senzacija.
Iritacija- izlaganje živog tkiva spoljašnjim i unutrašnjim stimulansima.
Reakcija- promjena (intenziviranje ili slabljenje) aktivnosti tijela ili njegovih komponenti kao odgovor na iritaciju.
refleksna reakcija- reakcija (ili proces) u tijelu (sistemu, organu, tkivu, ćeliji) uzrokovana refleksom.
Reflex- pojava ili promjena funkcionalne aktivnosti organa, tkiva ili cijelog organizma, koja se odvija uz učešće centralnog nervnog sistema kao odgovor na iritaciju nervnih završetaka (receptora).
Ekscitabilnost- sposobnost živih ćelija da percipiraju promjene spoljašnje okruženje i reagovati na ove promene odgovorom ekscitacije. Što je niži prag jačine stimulusa, to je veća ekscitabilnost, i obrnuto.
Uzbuđenje- aktivan fiziološki proces kojim neke žive ćelije (nervne, mišićne, žljezdane) odgovaraju na vanjske utjecaje.
Ekscitabilna tkiva- tkanine sposobne da reaguju na prekoračenje spoljašnjih uticaja prag ekscitacije, da se iz stanja fiziološkog mirovanja pređe u stanje uzbuđenja. U principu, sve žive ćelije imaju određeni stepen ekscitabilnosti, ali u fiziologiji je uobičajeno da se ova tkiva nazivaju nervnim, mišićnim i žlezdanim.
fiziologija ćelije
Ćelijske membrane.
Predavanje 2
transmembranski transport.
membranski potencijal.
Razlika u električnom potencijalu (u voltima ili mV) između tekućine na jednoj strani membrane i tekućine na drugoj strani naziva se membranski potencijal(MP) i označava se Vm. Magnituda magnetnog polja živih ćelija obično je od -30 do -100 mV, a sva ta razlika potencijala stvara se u područjima koja su direktno uz ćelijsku membranu sa obe strane. Smanjenje MF vrijednosti se naziva depolarizacija, povećati - hiperpolarizacija, vraćanje originalne vrijednosti nakon depolarizacije - repolarizacija. Membranski potencijal postoji u svim ćelijama, ali u ekscitabilnim tkivima (nervno, mišićno, žljezdano), membranski potencijal ili kako se još naziva u ovim tkivima, membranski potencijal mirovanja, igra ključnu ulogu u realizaciji njihovih fizioloških funkcija. Potencijal membrane je zbog dva glavna svojstva svih eukariotskih ćelija: 1) asimetrična distribucija jona između ekstra- i intracelularne tečnosti, podržana metaboličkim procesima; 2) Selektivna permeabilnost jonskih kanala ćelijskih membrana. Da biste razumjeli kako nastaje MF, zamislite da je određena posuda podijeljena na dva odjeljka membranom koja je propusna samo za jone kalija. Neka prvi odjeljak sadrži 0,1 M, a drugi 0,01 M rastvor KCl. Budući da je koncentracija kalijevih jona (K +) u prvom odjeljku 10 puta veća nego u drugom, tada će u početnom trenutku na svakih 10 K+ iona koji difundiraju iz odjeljka 1 u drugi odjeljak, doći će jedan ion koji difundira u obrnuti smjer. Budući da kloridni anjoni (Cl-) ne mogu proći kroz membranu zajedno s kalijevim kationima, u drugom odjeljku će se formirati višak pozitivno nabijenih jona i, naprotiv, višak Cl- jona će se pojaviti u odjeljku 1. Kao rezultat toga, postoji transmembranska razlika potencijala, što onemogućava dalju difuziju K+ u drugi pretinac, jer za to treba da savladaju privlačenje negativnih Cl- iona u trenutku ulaska u membranu iz odeljka 1 i odbijanje sličnih jona na izlazu iz membrane u odjeljak 2. Dakle, za svaki ion K+ koji u ovom trenutku prolazi kroz membranu djeluju dvije sile - hemijski koncentracijski gradijent (ili razlika hemijskog potencijala), koji doprinosi tranziciji kalijevih jona iz prvog odjeljka u drugi, i električnu potencijalnu razliku, tjerajući K+ ione da se kreću u suprotnom smjeru. Nakon što se ove dvije sile izbalansiraju, broj K+ jona koji se kreću iz odjeljka 1 u odjeljak 2 i obrnuto postaje jednak, elektrohemijska ravnoteža. Transmembranska razlika potencijala koja odgovara takvom stanju naziva se ravnotežni potencijal, u ovom konkretnom slučaju, ravnotežni potencijal za kalijeve ione ( Ek). Krajem 19. stoljeća Walter Nernst je ustanovio da ravnotežni potencijal zavisi od apsolutne temperature, valencije difuznog jona i omjera koncentracija ovog jona prema različite strane membrane:
gdje ex- ravnotežni potencijal za X ion, R- univerzalna plinska konstanta = 1,987 cal/(mol deg), T je apsolutna temperatura u stepenima Kelvina, F- Faradejev broj = 23060 cal/in, Z je naelektrisanje prenesenog jona, [X]1 i [x]2- koncentracija jona u odjeljcima 1 i 2.
Ako idemo od prirodnog logaritma do decimalnog logaritma, tada se za temperaturu od 18 °C i monovalentni ion Nernstova jednadžba može napisati na sljedeći način:
Ex= 0,058 lg
Pomoću Nernstove jednadžbe izračunavamo potencijal ravnoteže kalija za zamišljenu ćeliju, uz pretpostavku da je ekstracelularna koncentracija kalija [K + ]n = 0,01 M, a unutarćelijska je [K + ]v = 0,1 M:
Ek = 0,058 lg = 0,058 lg = 0,058 (-1) = -0,058 ‚= -58 mV
U ovom slučaju, Ek je negativan, budući da će joni kalija napustiti hipotetičku ćeliju, negativno nabijajući citoplazmatski sloj koji se nalazi uz unutrašnju stranu membrane. Budući da postoji samo jedan difuzni jon u ovom hipotetičkom sistemu, potencijal ravnoteže kalijuma će biti jednak potencijalu membrane ( Ek \u003d Vm).
Gore navedeni mehanizam je također odgovoran za formiranje membranskog potencijala u stvarnim stanicama, ali za razliku od razmatranog pojednostavljenog sistema, u kojem bi samo jedan ion mogao difundirati kroz "idealnu" membranu, prave ćelijske membrane propuštaju sve anorganske jone. jedno ili drugo. Međutim, što je membrana manje propusna za bilo koji ion, to ima manje utjecaja na magnetsko polje. S obzirom na ovu okolnost, Goldman je 1943. predložena je jednadžba za izračunavanje MF vrijednosti stvarnih ćelija, uzimajući u obzir koncentracije i relativnu permeabilnost svih difuznih jona kroz plazma membranu:
Vm = 0,058 lg
Koristeći metodu obilježenih izotopa, Richard Keynes je 1954. godine odredio permeabilnost mišićnih stanica žabe za osnovne ione. Pokazalo se da je permeabilnost za natrijum oko 100 puta manja nego za kalij, a Cl-ion ne daje značajan doprinos stvaranju MP u mišićnim ćelijama. Stoga se za membrane mišićnih ćelija Goldmanova jednačina može napisati u sljedećem pojednostavljenom obliku:
Vm = 0,058 lg
Vm = 0,058 lg
Studije pomoću mikroelektroda umetnutih u ćelije pokazale su da se potencijal mirovanja ćelija skeletnih mišića žabe kreće od -90 do -100 mV. Ovako dobro slaganje eksperimentalnih i teorijskih podataka potvrđuje da je potencijal mirovanja određen difuzijskim tokovima anorganskih iona. Istovremeno, u stvarnim ćelijama membranski potencijal je blizak ravnotežnom potencijalu jona, koji karakteriše maksimalna transmembranska permeabilnost, odnosno ravnotežni potencijal jona kalijuma.
Uloga kalcijuma
Predavanje 3
Osnove bioenergije.
Hemijska energija, karakteristike njenog nastanka i korišćenja u živim sistemima.
Bioenergetika proučava molekularne mehanizme pretvaranja energije u živim ćelijama. Redukovana organska jedinjenja, koja služe kao izvor energije živim organizmima, oksidiraju se do CO 2 i H 2 O, a oslobođena energija se koristi za sintezu ATP-a i obavljanje različitih zadataka.
Prije nego što pređemo na razmatranje specifičnih bioenergetskih procesa u ćeliji, podsjetimo se da, u skladu s drugim zakonom termodinamike, entropija svakog zatvorenog sistema može samo rasti s vremenom. Entropija je termodinamička kategorija koja karakteriše stepen nereda u sistemu. Dakle, povećanje entropija, zapravo dolazi do povećanja stepena poremećenosti sistema, tj. prelazak iz uređenije strukture u manje uređenu i smanjenje udjela korisna energija(tj. energija sa kojom se može obaviti rad). Drugim riječima, iz drugog zakona termodinamike slijedi da svakom izolovanom sistemu, stepen poremećenosti se može samo povećati, a količina korisne (slobodne) energije može smanjiti. Istovremeno, živi organizmi, kako se razvijaju od oplođenog jajeta do stanja odrasla osoba, stalno komplikuju njihovu strukturu. Čini se da drugi zakon termodinamike nije ispunjen u živim sistemima. Međutim, treba imati na umu da se ovaj zakon odnosi na zatvoreni sistem, a živi organizam je otvoreni sistem, koji karakteriše stalna razmena materije i energije sa okolinom i održavanje relativno niskog nivoa entropije povećanjem to u okruženju. Sposobnost izvlačenja korisne energije iz okruženje je jedno od onih izvanrednih svojstava koja razlikuju žive organizme od nežive prirode. Konzumacijom biljne ili životinjske hrane, živi organizam pretvara visokoorganizirane molekule sadržane u njemu u CO 2, H 2 O i spojeve niske molekularne težine koji sadrže dušik, kojih ima mnogo više. visoki nivo entropije, a rezultirajuća energija se koristi, uključujući razvoj i rast vlastitog organizma, praćen smanjenjem entropije. Primarni izvor energije na Zemlji za većinu organizama je sunce, čija se entropija stalno povećava i samo neznatan dio energije koja se u ovom slučaju oslobađa biosfera hvata i koriste biljke i neke bakterije u reakcijama fotosinteze, a ostalo je nepovratno izgubljeno u Univerzumu. Čak i nedavno otkriveno bogat život duboko na dnu okeana u području okeanskih pukotina na tzv. "Crni pušači" zavise od sunca jer je aerobno, a kiseonik se u atmosferu i okean uzima samo procesom fotosinteze.
U procesu evolucije u tijelu životinja i, naravno, nastalo je mnogo načina dobivanja, transporta i korištenja energije, a u velikoj većini njih se za akumulaciju kemijske energije koriste posebna jedinjenja - fosfageni, koji u svojoj strukturi imaju tzv visokoenergetski ili makroergijski veza. Adenozin-5"-trifosfat (ATP) je najčešći i najvažniji predstavnik ove grupe spojeva. Pored ATP-a, niz drugih organskih polifosfata ima visokoenergetske veze, na primjer, GTP, CTP. Simbol ~ se koristi za označavanje makroergijskih veza. Dakle, ATP molekul je skraćeno kao A-R-F~F~F(ALI-adenin, R-riboza). Neke bakterije čak pohranjuju energiju u obliku anorganskog polimetafosfata, linearnog polimera neodređene dužine.
Pri pH=7,0, ATP je anjon sa visokim nabojem. Trifosfatna grupa sadrži 4 OH grupe sposobne za jonizaciju. 3 od njih imaju nisku pK vrijednost i potpuno se disociraju pri pH=7, četvrta grupa se disocira za 75%. Visoka koncentracija negativnih naboja važan je faktor u visokoenergetskoj prirodi ATP-a. Ovi naboji su blizu jedan drugom i između njih postoji jaka odbojnost. Sa hidrolitičkim cijepanjem terminalne fosfatne grupe, odbojne sile slabe. Nastali proizvodi nemaju tendenciju ponovnog spajanja. Druga važna karakteristika koja određuje veliku negativnu vrijednost Δ G" Hidroliza ATP-a leži u činjenici da su oba produkta girolize stabilizirana konjugacijom. Međutim, ATP se gotovo nikada ne nalazi u ćelijama u obliku slobodnih jona, već češće u obliku kompleksa sa Mg ili Mn ionima.
Koncept energije fosfatne veze, koji koriste biohemičari, ne treba miješati s konceptom energije veze, što u fizičkoj hemiji znači energiju potrebnu za prekid veze između 2 atoma u molekuli. Pod energijom makroergične fosfatne veze podrazumijeva se razlika između slobodne energije polaznih supstanci i slobodne energije produkta reakcije, tokom koje se fosforilirano jedinjenje hidrolizira da nastane anorganski fosfat.
Transfer terminalne fosfatne grupe ATP na vodi uz ispuštanje slobodna energija, približno dvostruko veći nego kod hidrolize konvencionalne fosfoesterske veze, na primjer, u adenozin monofosfatu. Besplatna energija karakterizira onaj dio ukupne energije sadržane u tvari, uz pomoć kojeg se može obaviti rad izotermni i izobarični uslovi, tj. pri konstantnoj temperaturi i pritisku, naime takvi uslovi su tipični za biološke sisteme. Promjena slobodna energija mjereno u kcal/mol i označava se sa Δ G" ako se proces odvija u standardnim uslovima (rN=7, T=25 ºS, supstrati i produkti reakcije se uzimaju u istoj koncentraciji jednakoj 1 mol/l) i simbol Δ G za reakcije u specifičnim uslovima stvarnih bioloških sistema. U slučaju hidrolize ATP-a, Δ G"\u003d -7 kcal / mol i Δ G\u003d -11-13 kcal / mol Živi organizmi koriste energiju ATP-a za obavljanje mehaničkog rada mišića, stvaranje gradijenata koncentracije i transmembranskih električnih potencijala.
Konkretni mehanizmi koji obezbeđuju tok ovih procesa biće izloženi u narednim predavanjima, a sada ćemo razmotriti opšte principe koji omogućavaju korišćenje energije ATP-a za obavljanje hemijskog rada. Zamislite tipičan biosintetik, tj. anabolički proces (anabolizam od grčkog uspona, oni procesi sinteze, izgradnja strukturnih dijelova ćelije, suprotni katabolizmu), u kojima se dva monomera -X i Y- moraju spojiti jedan s drugim u reakciji dehidracije (koja se naziva i kondenzacija), praćeno ispuštanjem vode:
X-OH + Y-H X-Y + H 2 O + Δ G (3 kcal/mol)
Većina ovih reakcija je endergonic(ne treba ih brkati sa endotermnim) i praćeni su povećanjem slobodne energije za Δ G, a pokretačka snaga bilo koje kemijske reakcije je smanjenje slobodna energija i spontano teče exergonic reakcija i stoga katabolička reverzna reakcija (nazvana hidroliza) je gotovo uvijek poželjna, jer u ovom slučaju oslobađanje slobodna energija jednaka po apsolutnoj vrijednosti njegovoj potrošnji u reakciji kondenzacije. Stoga se u ćeliji takve reakcije odvijaju u nekoliko faza, zbog čega je energetski nepovoljna sinteza povezana sa snažnom exergonic reakcija koja uključuje ATP. Mehanizam uparivanja se implementira na sljedeći način. Reakcija fuzije koja je gore navedena i praćena povećanjem slobodne energije (pretpostavimo Δ G" \u003d 3 kcal / mol), kojoj prethodi fosforilacija jednog od supstrata molekulom ATP.
A-R-F~F~F + HON → A-R-F~F + X-O-F - Δ G = 4 kcal/mol
Δ G" ova reakcija je mnogo manja 4 kcal/mol) nego reakcije hidrolize ATP i transfer fosfatne grupe u vodu (oko 11 kcal/mol). Ostatak energije ( 7 kcal/mol) se akumulira u aktiviranoj molekuli supstrata, koja je uključena u naknadnu sintetičku reakciju. Međutim, sada je slobodna energija polaznih supstanci veća od slobodne energije produkta reakcije (7 - 3 = 4 kcal/mol) i reakcija se odvija s lijeva na desno, tj. u smjeru fuzije, praćeno smanjenjem slobodne energije:
X-O-F + Y-H → X-Y + F n - Δ G = (4 kcal/mol)
Dakle, zbog energije pohranjene u ATP molekulu endergonična reakcija zamijenjen sa dva konjugata exergonic reakcije.
Drugi način korištenja ATP-a za obavljanje kemijskog rada je kroz sekvencijalno konjugirane, ili kako ih zovu, tandemske reakcije. U ovom slučaju endergonična reakcija nastavlja zahvaljujući sledećem exergonic reakcije. Mogućnost nastanka tandemskih reakcija je zbog činjenice da povećanje slobodna energija ne isključuje nastanak hemijske reakcije, već samo ukazuje da je vrijednost konstante ravnoteže takve reakcije manja od jedinice, tj. ako postoji reakcija:
A + B C + D + ΔG" 1 (1)
onda do ekv= < 1
i, posljedično, ravnoteža reakcije se pomjera ulijevo, a taj pomak je utoliko izraženiji što je veća apsolutna vrijednost Δ G". Međutim, ako nakon reakcije 1 slijedi reakcija 2
S +X CX- Δ G" 2 (2)
a za ovu reakciju vrijednost Δ G" je jako negativan, tada će se intermedijar reakcije C potrošiti, pomjerajući ravnotežu reakcije 1 udesno. U slučaju da je ukupna slobodna energija dvije reakcije negativna, one će se nastaviti u smjeru potrošnje supstrata A i B i formiranja proizvoda CX.
Sada razmotrimo gdje su u ćeliji lokalizirani mehanizmi za dobivanje energije i prijenos fosfagena iz "nenabijenog" oblika (na primjer, ADP) u "nabijeni" (ATP) i kako funkcioniraju.
Ciklus trikarboksilne kiseline
U većinu ćelija i tkiva aerobnih organizama ulazi pirogrožđana kiselina nastala kao rezultat glikolize. mitohondrijski matriks, gdje je dekarboksiliran, tj. od njega se odvaja molekul ugljičnog dioksida, a preostala dva ugljika acetil grupa povezuje sa koenzim A (COA), formiranje acetil-CoA. Energija koja se oslobađa kao rezultat ove reakcije troši se na prijenos posebne molekule nosača vodika iz oksidiranog u reducirani oblik. Nosač vodika naziva se nikotinamid adenin dinukleotid i skraćeno je NAD (IZNAD) ili NAD+ (oksidirani oblik) i NADH (NADH) ili NADH + H+ (redukovani oblik). Dakle, NAD+ molekuli i oni slični njemu po strukturi i funkciji flavin adenin dinukleotid, skraćeno FAD, kao i fosfageni, oni su u stanju skladištiti i prenositi energiju, ali ne u obliku visokoenergetske fosfatne veze, već u obliku aktiviranih atoma vodika. Acetil-CoA formiran iz pirogrožđane kiseline podleže daljoj oksidaciji u ciklus trikarboksilne kiseline. Kao rezultat prve reakcije ciklusa (sl.) iz acetil-C i oksalooctene kiseline (u vodenom mediju postoji u obliku jona - oksaloacetata) koji sadrži 4 atoma ugljika uz učešće enzima citrat sintaze, trikarboksilna, šestougljenična limunova kiselina(citrat). Stoga se ponekad naziva ciklus trikarboksilne kiseline ciklus limunske kiseline. Limunska kiselina se zatim pretvara u isolimon(izocitrat). Kao rezultat sljedeće reakcije dekarboksilacije, molekula izocitritne kiseline sa šest ugljika pretvara se u petougljičnu kiselinu α-oksoglutarna kiselina(α-oksoglutarat) i NAD+ se reducira na NADH. Nadalje, α-oksoglutarna kiselina se dekarboksilira u amber(sukcinat). Pritom se oslobođena energija ne raspršuje u obliku topline, već se troši na redukciju druge NAD+ molekule u NADH i formiranje međuproizvoda s visokoenergetskom fosfatnom vezom, koji osigurava fosforilaciju. gvanozin difosfat(BDP)in gvanozin trifosfat(GTP).
Kao rezultat sljedeće tri reakcije, sukcinska kiselina se sukcesivno pretvara u fumarna kiselina (fumarat), jabučna kiselina(malat) i oksaloacetat i obnavlja se pomoću molekula FAD i NAD.
Sa stanovišta bioenergije, na svakom koraku ciklusa uklanjaju se 2 atoma ugljika koji su ušli u ciklus kao acetil-CoA u obliku CO 2 i na ulazu se stvara 8[H] + 2[H] - ovo je aktivirani izvor energije vodika.
Dakle, enzimske reakcije ciklusa trikarboksilne kiseline odvijaju se na način da je supstrat prve reakcije oksalosirćetne kiseline je također proizvod posljednje osme reakcije, tj. se regeneriše nakon svakog okretanja ciklusa i proces počinje iznova. Kao rezultat elementarnog ciklusa, dva atoma ugljika prisutna u acetil-CoA se pretvaraju u CO 2 . Cikličnu prirodu sekvence reakcije oksidacije acetil-CoA i njenih glavnih faza utvrdio je 1937. godine engleski biohemičar njemačkog porijekla, dobitnik Nobelove nagrade (1953.) Hans Krebs. Stoga proces oksidacije acetil-CoA ima drugo ime - Krebsov ciklus. Zbog činjenice da se oksidacija odvija postepeno, nizom relativno malih promjena slobodne energije, oslobođena energija se ne raspršuje u obliku beskorisne topline, već se troši na redukciju tri NAD molekule u NADH i jedne FAD molekule u FADH i formiranje visokoenergetske fosfatne veze. Ova veza nastaje fosforilacijom gvanozin difosfata u gvanozin trifosfat. Potonji lako izmjenjuje fosfatnu grupu sa ADP-om da bi formirao ATP.
Treba imati na umu da su, uz glikolizu, i drugi biohemijski procesi koji se odvijaju u ćelijama, posebno oksidacija aminokiselina i masnih kiselina (kao i etanola), izvor acetil-CoA za oksidaciju u TCA. Četiri molekula NADH, uključujući onaj koji nastaje tokom oksidacije piruvata u acetil-CoA, i molekula FADH ulaze u respiratorni lanac, gdje se atomi vodika koje nose oksidiraju molekularnim kisikom u vodu. Energija oslobođena u ovom slučaju troši se na sintezu ATP-a iz ADP-a i anorganskog fosfata kao rezultat najsloženijeg, uglavnom nejasnog procesa, tzv. oksidativna fosforilacija.
Predavanje 4
Predavanje 5
Predavanje 6
Lizozomi: struktura i načini formiranja u ćeliji, klasifikacija.
Lizosomi su heterogena (heterogena) grupa citoplazmatskih vakuolnih struktura veličine 1-3 μm, čija je karakteristična karakteristika prisustvo kiselog okruženja i velikog broja različitih hidrolaza - enzima sposobnih za cijepanje (hidrolizaciju) glavnog vrste makromolekula. Prisustvo hidrolaza u lizosomima određuje njihovu glavnu funkciju u ćeliji - cijepanje makromolekula i većih formacija, koje ulaze u ćeliju iz ekstracelularnog prostora i one intracelularnog porijekla. Zanimljivo je da biljke nemaju lizozome.
Poznata su tri moguća puta za formiranje lizosoma u ćeliji. U svakom slučaju, formiraju se morfološki različite formacije, cijepajući materijal iz različitih izvora.
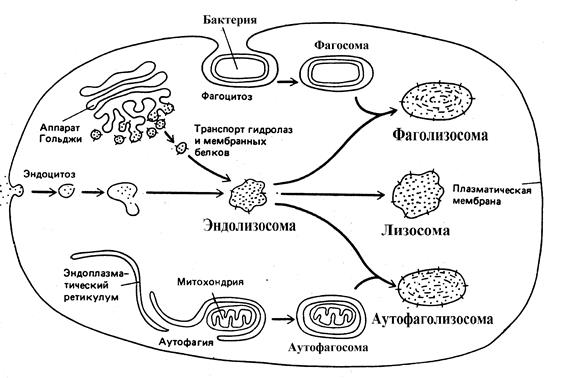 Rice. 6 Načini stvaranja lizosoma u ćeliji
Rice. 6 Načini stvaranja lizosoma u ćeliji
U prvom slučaju materijal koji se može cijepati - proteini, polinukleotidi ili polisaharidi - ulazi u ćeliju putem endocitoza. Tokom ovog procesa, molekule koje su dovoljno velike i ne mogu prodrijeti kroz membrane postupno su okružene malim dijelom plazmaleme, koji prvo invaginira (invaginira), a zatim se odcjepljuje u ćeliju, formirajući vezikulu koja sadrži materijal koji je stanica uhvatila. . Mjehurići nastali kao rezultat endocitoze nazivaju se endosomi. Kako se endosom kreće od ćelijske membrane u unutrašnjost ćelije, on stalno stupa u interakciju s transportnim vezikulama koje isporučuju iz trans- površina Golgijevog aparata hidrolitički enzimi i membranski proteini, koji se pretvaraju u endolizozom. Proces formiranja i transformacije endosoma traje oko 15 minuta i praćen je zakiseljavanjem unutrašnje sredine zbog pumpanja H+ jona iz citosola u endosom pomoću ATP zavisne protonske pumpe, koja funkcioniše kao ATP-aza. unutrašnje mitohondrijalne membrane.
Drugi put za formiranje lizosoma se zove autofagija. U procesu autofagije uništavaju se istrošeni dijelovi same ćelije. Poznato je, na primjer, da je u ćelijama jetre prosječan životni vijek jedne mitohondrije oko 10 dana, nakon čega se mora iskoristiti u lizozomima. Na elektronskim mikrosnimcima normalnih ćelija mogu se vidjeti lizozomi koji sadrže mitohondrije u različitim fazama degradacije. Gore je već napomenuto da se autofagijom uklanja višak glatkog ER iz ćelija jetre nakon prestanka unosa i izlučivanja ksenobiotika - indukatora iz organizma. Čini se da proces autofagije počinje okruživanjem organele membranama dostavljenim iz ER, što rezultira formiranjem autofagozom. Tada se vjeruje da se autofagosom spaja sa endolizozomom, formirajući autofagolizosom, u kojem dolazi do procesa degradacije ER fragmenta ili druge organele.
Treći put za formiranje lizosoma dostupan je samo u ćelijama specijalizovanim za fagocitoza velike čestice i mikroorganizmi. Takve fagocitne ćelije, a tu spadaju i krvne ćelije - neutrofili i monociti, mogu apsorbovati velike objekte iz ekstracelularnog prostora, formirajući fagozomi. Fagozom tada postaje fagolizozom na isti način kao i autofagosom, tj. spajanje sa endolizozomom.
Endosomi, autofagosomi i fagozomi se često nazivaju uobičajenim terminom - prelizozomi, a endolizosomi, autofagolizozomi i fagolizosomi se nazivaju lizosomi. U zrelim lizosomima, apsorbirani materijal se razgrađuje do pojedinačnih molekula, kao što su aminokiseline, koje ulaze u citosol i učestvuju u kasnijim biohemijskim transformacijama. Fragmenti sopstvene plazma membrane nisu izloženi hidrolazama i vraćaju se nazad u plazma membranu uz pomoć transportnih vezikula, čak i pre konačnog formiranja lizosoma. Nesvarljivi proizvodi ostaju i akumuliraju se u lizosomima, koji gube hidrolitičke enzime i pretvaraju se u postlizozomi ili zaostala tijela. S godinama, u ljudskim i životinjskim ćelijama, broj zaostalih tijela sadrži veliki broj lipofuscin ili pigmenta za starenje.
Lipofuscin je biopolimer različite prirode, koji nije podložan daljem cijepanju jer hemijske veze između pojedinačnih monomera nisu nastali u normalnim biohemijskim reakcijama, već kao rezultat spontanih oksidativnih procesa, uglavnom slobodnih radikala. Razne bolesti, izloženost zračenju i drugi negativni čimbenici okoline ubrzavaju nakupljanje pigmenta koji stari.
Ćelijsko jezgro i genom
Predavanje 7
Predavanje 1
Uvod u kurs
Predmet i zadaci fiziologije
Fiziologija (od grčkih reči: physis - priroda, logos - učenje, nauka) je nauka o funkcijama kako tela u celini, tako i njegovih pojedinačnih komponenti: organa, tkiva, ćelija; procesi i mehanizmi koji omogućavaju implementaciju i regulaciju ovih funkcija i osiguravaju vitalnu aktivnost ljudi i životinja u njihovoj interakciji sa okolinom. Zadatak fiziologije je da uspostavi fizičke i hemijske osnove životnih procesa na svim nivoima organizacije živih sistema od subćelijskog do nivoa celog organizma. U vezi sa složenošću predmeta izučavanja, fiziologija je podijeljena na nekoliko cjelina. To uključuje: opštu i ćelijsku fiziologiju, čiji je zadatak proučavanje obrazaca osnovnih životnih procesa i mehanizama karakterističnih za sve funkcionalne i strukturne sisteme tela, i privatnu fiziologiju, koja se dalje deli na fiziologiju pojedinih organa: srce, bubrezi, jetra itd.; fiziologija funkcionalnih sistema: disanje, cirkulacija, probava, endokrini sistem, razvojna fiziologija, neurofiziologija i fiziologija ponašanja, senzorna fiziologija. Postoji i primijenjena fiziologija: fiziologija sporta, rada, ishrane, svemira i podvodne fiziologije.
Koji je praktični značaj fiziologije? Zašto je važno znati kako tijelo funkcionira? Najočigledniji razlog je taj što je fiziologija, a posebno proučavanje funkcionisanja ljudskog tijela, naučna osnova medicine i farmaceutske hemije. U svim prethodnim fazama razvoja civilizacije pristup liječenju bolesti bio je gotovo u potpunosti empirijski, tj. svelo se na pokušaje i greške. Kao rezultat toga, postepeno se uočavalo da određeni lijekovi pomažu kod određenih bolesti – bilo da se radi o hemijskim ili biljnim preparatima, mineralnoj vodi i toplim kupkama, akupunkturi, pa čak i metodama psihološkog utjecaja koje su bile široko korištene u narodne medicine. Iscjeljujuće djelovanje mnogih moderne droge je prvi put otkriven čisto empirijski (na primjer, nitroglicerin, koji je Nobel koristio za proizvodnju dinamita, istovremeno izazivajući ozbiljan pad krvnog tlaka kod radnika, što često dovodi do smrti), a glavni mehanizmi koji su u osnovi ljekovitog djelovanja brojnih njih još uvijek su ostaju nepoznati. Međutim, kako se naše razumijevanje mehanizama funkcionisanja i patologije organa i tkiva razvija i produbljuje, uloga čisto empirijskih nalaza se smanjuje i postaje moguće razviti učinkovite, na dokazima zasnovane metode za liječenje ljudskih bolesti, ciljano stvaraju
1.1. Opšti pogled o fiziologiji
Main pitanja: Organizam i stanište. Razumijevanje fiziologije. Fiziološka funkcija i njene multiparametarske karakteristike. Homeostaza, konstante opuštanja i njihovi tipovi. Metode istraživanja u fiziologiji. Akutni i kronični eksperimenti. Analitički i sistemski pristupi proučavanju fizioloških funkcija. Teorija funkcionalnih sistema prema P.K. Anokhin.
Ljudski organizam- holistički, samoregulirajući životni sistem sposoban osjećati, razmišljati, aktivno se ciljano kretati, prilagođavati okolini ili je prilagođavati svojim biološkim i društvenim potrebama.
Biološko stanište nazivaju ukupnost prirodnih uslova neophodnih za normalno postojanje živih tela.
Postoje spoljašnje i unutrašnje biološke sredine. Eksterno okruženje je kompleks prirodnih faktora koji su izvan tijela, ali neophodni za održavanje njegove vitalne aktivnosti.
Ukupnost bioloških tečnosti koje kupaju ćelije tela su unutrašnje sredine tela . Tečnosti unutrašnje sredine tela - krv, limfa, međućelijska tečnost i druge, spoljašnje su okruženje za njegove ćelije.
živa ćelija je elementarna strukturna i funkcionalna jedinica tijela. Objedinjavanje ćelija koje osiguravaju obavljanje određenog specifičnog zadatka dovelo je u procesu evolucije do formiranja živa tkiva – pokrivač (epitelne), masno, kost, vezivni, žljezdani, mišićav i nervozan. Živa tkiva su tijela- srce, pluća, jetra, bubrezi i drugi, koji osiguravaju provedbu složenih diferenciranih funkcija usmjerenih na održavanje vitalne aktivnosti tijela.
Nauka o vitalnoj aktivnosti zdravog organizma u njegovoj interakciji sa spoljašnjom sredinom, koja proučava funkcije ćelija, tkiva, organa, sistema organa i tela u celini, kao i mehanizme njihove regulacije, naziva se fiziologija (grčki "physis" - priroda, "logos" - nauka).
Postoje tri glavna zadatka fiziologije:
1) proučavanje objektivnih obrazaca toka fizioloških funkcija (šta se dešava?),
2) rasvetljavanje mehanizama za realizaciju fizioloških funkcija (kako?),
3) utvrđivanje namjene fizioloških funkcija (zašto?).
Fiziološka funkcija(latinski "functio" - aktivnost) - specifičan oblik aktivnosti tijela, koji se završava postizanjem određenog rezultata korisnog za njega, omogućavajući mu da se prilagodi uvjetima okoline.
Dakle, glavna funkcija srca je da pumpa krv u vaskularni krevet, a gastrointestinalnog trakta osigurava opskrbu krvi hranjivim tvarima. Većina organa i sistema obavlja nekoliko funkcija. Na primjer, glavna funkcija bubrega je izlučivanje krajnjih produkata metabolizma, ali su također uključeni u regulaciju krvnog tlaka, osmotskog tlaka i ravnoteže elektrolita tjelesnih tekućina.
Fiziološke funkcije su manifestacija vitalne aktivnosti organizma i karakterišu ih određeni znaci - parametri. Istu fiziološku funkciju može karakterizirati nekoliko parametara, među kojima su:
1) intenzitet fiziološke funkcije,
2) ekstenzivnost,
3) moć,
4) faktor efikasnosti (COP),
5) vremenske karakteristike,
6) bioritmovi.
Intenzitet karakteriše nivo intenziteta fiziološke funkcije. Ovi parametri se izražavaju u apsolutnoj vrijednosti kvalitativnih manifestacija fizioloških funkcija - fiziološke konstante, tj. indikatori unutrašnjeg okruženja tijela (krvni tlak, tjelesna temperatura, koncentracija glukoze u krvi i dr.).
Ekstenzivnost pokazuje interakcijom kojih procesa se postiže intenzivan parametar. Dakle, intenzivan parametar funkcije termoregulacije - temperatura ljudskog tijela, predodređen je ekstenzivnošću dva različito usmjerena procesa - proizvodnje topline i prijenosa topline.
Snaga je rad koji organizam obavi u jedinici vremena.
efikasnost(efikasnost) je omjer energije koju tijelo potroši za obavljanje korisnog rada prema cijeloj energiji koja je utrošena u ovom slučaju.
Tajming uključiti brzina tok fizioloških procesa i njihov ubrzanje. konačno, bioritmovi - To su periodične ciklično ponavljajuće promjene fizioloških funkcija. Prema trajanju perioda, razlikuju se tri glavne vrste bioritma:
1) ultradijanski, sa frekvencijom od djelića sekunde do 20 sati,
2) cirkadijanski (danonoćno) - od 20 do 28 sati,
3) infradijanski, koji uključuju sedmično, period, sezonski i godišnji bioritmovi.
Parametri fiziološke funkcije mogu se promijeniti pod utjecajem regulatorni mehanizmi. Ispod biološka regulacija razumjeti takvu kontroliranu promjenu fiziološke funkcije, koja ima za cilj osiguranje određene aktivnosti živog sistema ili njegovu otpornost na djelovanje remetilačkih faktora.
Unatoč činjenici da tijelo neprestano razmjenjuje materiju, energiju i informacije sa okolinom, svako živo biće je u stanju održati nepromjenjivost i stabilnost svojih unutrašnjih vitalnih parametara. Relativna konstantnost unutrašnjeg okruženja tela i stabilnost njegovih fizioloških funkcija se nazivaju homeostaza.
Homeostazu karakteriše ukupnost različitih fizioloških konstanti. Takve konstante održava živi sistem blizu nivoa koji unaprijed određuje optimalnu ćeliju metabolizam - skup procesa transformacije supstanci i energije koji osiguravaju vitalnu aktivnost organizma i njegov odnos sa vanjskim okruženjem. Stoga je homeostaza apsolutno neophodan uslov za život.
Organizam je otvoreni termodinamički sistem koji neprestano razmjenjuje materiju, energiju i informacije sa okolinom. Stoga fiziološki parametri ne mogu biti apsolutno stabilni. Konstante koje se mijenjaju u granicama koje osiguravaju biološki optimum života i normalan tok metabolizma nazivaju se konstante opuštanja .
Prema amplitudi promjene, relaksacijske konstante se dijele na dva tipa - tvrd i plastika. Dozvoljena amplituda fluktuacije nivoa tvrde konstante mala. To uključuje fizičko-hemijske parametre unutrašnje sredine, posebno osmotski i onkotski pritisak krvne plazme, njenu kiselo-baznu ravnotežu. Plastične konstante karakteriše relativno velika dozvoljena amplituda fluktuacije njihove veličine. To uključuje takve fiziološke pokazatelje kao što su tjelesna temperatura, krvni tlak, otkucaji srca i disanje, protok krvi i mnogi drugi.
Za održavanje homeostaze tijelo mora:
1) konzumiraju i razlažu hranu na hranljive materije koje zadovoljavaju energetske i plastične potrebe ćelija, tkiva i organa,
2) apsorbuju kiseonik kako bi oksidirali hranljive materije, koji su izvor energije živog sistema,
3) ispuštanje u životnu sredinu nepotrebnih i štetnih metaboličkih produkata,
4) imati sposobnost kretanja kako bi uhvatio hranu, pobjegao od neprijatelja.
Ako pokazatelji homeostaze prelaze normalne raspone koji određuju optimalni tijek metabolizma, onda to dovodi do kršenja fizioloških funkcija, razvoja bolesti i smrti.
Proučiti indikatore unutrašnjeg okruženja tijela i njegovih fizioloških funkcija, metode kliničkog istraživanja i eksperimenti. Kliničke studije se provode na ljudima, a eksperimenti na životinjama.
U ranim fazama razvoja fiziologije bile su posebno popularne kirurške eksperimentalne tehnike: ekstirpacija uklanjanje dijela ili cijelog organa i transplantacija - transplantaciju ispitivanog organa u istom organizmu na novu lokaciju ili njegovo prenošenje u drugi organizam, nakon čega slijedi posmatranje i evidentiranje posljedica takvih intervencija.
Za proučavanje aktivnosti organa koji su nedostupni direktnom posmatranju koriste se metoda fistule . Sastoji se u brzom stvaranju poruke od unutrašnjeg organa sa spoljašnjim okruženjem. Varijacija ove tehnike može biti kateterizacija - uvođenje posebnih sintetičkih cijevi u kanale žlijezda ili u krvne žile.
Da bi se utvrdila zavisnost funkcije organa od uticaja nervnog sistema, koristi se tehnika denervacija . U tu svrhu se sijeku nervna vlakna koja inerviraju organ.
Sa pojavom moderne tehnologije, instrumentalne metode - elektrofiziološki, biohemijski, radiološki i mnoge druge, koje omogućavaju snimanje vitalnih procesa bez preliminarnih hirurških operacija, što omogućava proučavanje fizioloških funkcija ne samo kod životinja, već i kod ljudi.
U fiziologiji postoje dvije vrste eksperimenata koji su povezani s potrebom za izvođenjem hirurške intervencije: ljuto(vivisekcija) i hronično.
akutni eksperiment karakteriziraju:
1) odsustvo potrebe za održavanjem steriliteta u procesu vivisekcije,
2) sprovođenje istraživanja tokom ili neposredno nakon operacije,
3) eutanazija- ubijanje životinje tokom eksperimenta ili nakon njegovog završetka.
Akutni eksperiment je prilično grub upad istraživača u organizam. To je neophodno za početno prikupljanje podataka o funkcijama organa, tkiva i ćelija tijela. Metoda akutnog eksperimenta je metodološka osnova analitički pristup na proučavanje funkcija. Ovaj pristup karakteriše činjenica da su predmet proučavanja funkcije pojedinih ćelija, tkiva i organa ne uzimajući u obzir njihov međusobni odnos i sa okolinom.
hronični eksperiment karakteriziraju:
1) potreba održavanja sterilnosti tokom pripremnih hirurška operacija,
2) provođenje istraživanja tek nakon oporavka životinje,
3) ponovljeno proučavanje fizioloških funkcija organa ili organizma kao celine u uslovima što je moguće bližim prirodnim.
Metoda kroničnog eksperimenta je osnova sistemski pristup za proučavanje tjelesnih funkcija. Sistematski metodološki pristup karakteriše proučavanje fiziološke funkcije u odnosu na funkcionalno stanje organizma, kao celine, i uzimajući u obzir njegovu interakciju sa spoljašnjim okruženjem.
Osnivač sistemskog pristupa je ruski fiziolog, nobelovac I.P. Pavlov. Početkom 20. veka predložio je metod fiziološka hirurgija, koji omogućava proučavanje fizioloških funkcija u uslovima prirodnog ponašanja i po prvi put iznosi ideje o sistemska priroda aktivnosti tijela.
Skup organa koji obavljaju jednu funkciju je anatomski sistemi . Osoba ima devet anatomskih sistema:
1) muskuloskeletni , koji čini kostur tijela, osiguravajući kretanje njegovih dijelova jedan u odnosu na drugi i kretanje tijela u prostoru;
2) kardiovaskularni , obezbeđujući kretanje krvi i limfe kroz sudove;
3) respiratorni neophodan za isporuku kisika u tkiva i uklanjanje ugljičnog dioksida iz tijela;
4) digestivni dizajnirani za preradu hrane i transport rezultirajućih hranjivih tvari u krv ili limfu;
5) izlučivanje , koji osigurava uklanjanje metaboličkih proizvoda štetnih i nepotrebnih za tijelo;
6) endokrine , proizvodnju biološki aktivnih tvari - hormona uključenih u regulaciju fizioloških funkcija tijela;
7) seksualno , koji obavlja funkciju reprodukcije;
8) senzorni uključeni u percepciju signala iz vanjskog svijeta i unutrašnjeg okruženja tijela;
9) nervozan , koji ujedinjuje i reguliše aktivnost svih organa, osiguravajući interakciju organizma sa vanjskim okruženjem.
Privremeno povezivanje organa, tkiva i ćelija povezanih jednim zadatkom, čiji je cilj postizanje određenog rezultata vitalne aktivnosti korisne za tijelo, naziva se funkcionalni sistem .
Funkcionalni sistem(prema P.K. Anokhin) je složen dinamički, samoregulirajući i samoformirajući kompleks, koji se sastoji od centralnih i perifernih elemenata koji međusobno djeluju u procesu postizanja adaptivnog rezultata koji je koristan za tijelo i osigurava normalan metabolizam.
Funkcionalni sistem se sastoji od pet glavnih elemenata:
1) korisni adaptivni rezultat (PPR),
2) rezultat receptora,
3) obrnuta aferentacija,
4) nervni centar,
5) izvršni mehanizmi.
Kraj rada -
Ova tema pripada:
Udžbenik "OPŠTA FIZIOLOGIJA"
Supreme stručno obrazovanje... DRŽAVNA MEDICINSKA AKADEMIJA TVER... MINISTARSTVA ZDRAVLJA I SOCIJALNOG RAZVOJA RUSKOG FEDERACIJE...
Ako vam je potreban dodatni materijal na ovu temu, ili niste pronašli ono što ste tražili, preporučujemo da koristite pretragu u našoj bazi radova:
Šta ćemo sa primljenim materijalom:
Ako vam se ovaj materijal pokazao korisnim, možete ga spremiti na svoju stranicu na društvenim mrežama:
0001 Fiziološke reakcije živog organizma
Svaki živi organizam i sve njegove ćelije imaju razdražljivost, odnosno sposobnost da na uticaje okoline ili poremećaje u svom stanju reaguju promenom strukture, nastankom, jačanjem ili slabljenjem svoje snažne aktivnosti, koja je neraskidivo povezana sa kvalitativnim i kvantitativnim promenama u metabolizam i energiju. Promjene u građi i funkcijama tijela i njegovih stanica kao odgovor na različite utjecaje nazivaju se biološke reakcije, a utjecaji koji ih izazivaju nazivaju se podražaji ili podražaji. Pojam biološke reakcije je sve vrste aktivnosti odgovora organizma, njegovih organa i ćelija na različite uticaje. Reakcije ćelija se manifestuju u promeni oblika, strukture, procesa rasta i deobe, u stvaranju različitih hemijskih jedinjenja u njima, pretvaranju potencijalne energije u kinetičku (električnu, mehaničku, toplotnu, svetlosnu), performansama ćelija. jednog ili drugog rada (kretanje u prostoru, oslobađanje određenih supstanci, osmotski rad na koncentraciji određenih elektrolita u ćeliji). Reakcije cijelog organizma su izuzetno raznolike. U procesu njihovog sprovođenja menja se aktivnost mnogih organa i bezbroj ćelija, jer telo uvek reaguje na različite uticaje kao celina, kao jedan složen sistem. Stoga, iako se reakcije organizma odvijaju djelovanjem stanica, one se ne mogu svesti na reakcije pojedinih stanica. Ovo pokazuje opšte pravilo da se pravilnosti sistema ne mogu svesti na pravilnosti pojedinačnih elemenata koji čine sistem.
Iritacija Iritans za živu ćeliju ili organizam u cjelini može biti svaka promjena u vanjskoj sredini ili unutrašnje stanje organizam, ako je dovoljno velik, nastaje dovoljno brzo i traje dovoljno dugo. Sva beskonačna raznolikost mogućih iritansa ćelija i tkiva može se podijeliti u tri grupe: fizičke, fizičko-hemijske i hemijske. Fizički podražaji uključuju temperaturu, mehanički (udar, ubod, pritisak, kretanje u prostoru, ubrzanje, itd.), električne, svjetlosne i zvučne. Fizičko-hemijski stimulansi
su promjene osmotskog tlaka, aktivna reakcija medija, sastav elektrolita u koloidnom stanju. Hemijski iritansi uključuju mnoge tvari različitog sastava i svojstava koje mijenjaju metabolizam ili strukturu stanica. Hemijski iritansi koji mogu izazvati fiziološke reakcije su supstance hrane koje dolaze iz spoljašnje sredine, lekovi, otrovi, kao i mnoga hemijska jedinjenja koja nastaju u organizmu, kao što su hormoni, produkti metabolizma. Ćelijski podražaji koji izazivaju njihovu aktivnost su nervnih impulsa. Nervni impulsi, dolazeći duž nervnih vlakana od nervnih završetaka do centralnog nervnog sistema ili dolazeći iz njega u periferne organe – mišiće, žlezde, izazivaju promene u njihovom stanju i aktivnosti. Prema svom fiziološkom značaju, svi podražaji se dijele na adekvatne i neadekvatne. Adekvatni su oni podražaji koji djeluju na datu biološku strukturu u prirodnim uvjetima, čijem opažanju je posebno prilagođena i osjetljivosti na koju je izuzetno velika. Za štapiće i čunjeve mrežnjače adekvatan iritans su zraci vidljivog dela sunčevog spektra, za taktilne receptore kože - pritisak, za ukusne pupoljke jezika - razne hemikalije, za skeletne mišiće - nervni impulsi koji do njih teku duž motoričkih nerava. Neadekvatni su oni podražaji za čije opažanje data ćelija ili organ nije posebno prilagođen. Dakle, mišić se kontrahira kada je izložen kiselini ili lužini, električnoj struji, iznenadnom istezanju, mehaničkom udaru, brzom zagrijavanju itd. Ćelije su mnogo osjetljivije na svoje adekvatne podražaje nego na neadekvatne. Ovo je izraz funkcionalne adaptacije razvijene u procesu evolucije.
Ekscitabilnost Neke ćelije i tkiva (nervna, mišićna i žljezdana) su posebno prilagođena da izvode brze reakcije na iritaciju. Takve ćelije i tkiva nazivaju se ekscitabilnosti, a njihova sposobnost da na iritaciju reaguju ekscitacijom. Mjera ekscitabilnosti je minimalna snaga stimulusa koji izaziva ekscitaciju. Ovo
minimalna jačina iritacije naziva se prag iritacije. Što je veća minimalna snaga iritacije potrebna za izazivanje reakcija, to je veći prag iritacije, niža je razdražljivost, i obrnuto, što je niži prag iritacije, to je veća ekscitabilnost. U odnosu na različite podražaje, prag iritacije može biti različit. Posebno je visoka ekscitabilnost receptora u odnosu na adekvatne podražaje, na primjer, da bi se iritirala mirisna stanica, dovoljno je da na nju djeluje samo nekoliko molekula mirisne tvari.
Uzbuđenje Ekscitabilne stanice karakterizira specifičan oblik odgovora na djelovanje podražaja: u njima se odvija valoviti fiziološki proces - ekscitacija je složena biološka reakcija koja se manifestira u kombinaciji fizičkih, fizičko-kemijskih, kemijskih procesa i funkcionalnih promjena. Obavezni znak ekscitacije je promjena električnog stanja površinske ćelijske membrane. Kada su uzbuđene, ćelije prelaze iz stanja fiziološkog mirovanja u stanje fiziološke aktivnosti karakteristično za datu ćeliju: mišićno vlakno se skuplja, žljezdana ćelija luči tajnu.. U ekscitabilnoj ćeliji postoji konstantna razlika u električnim potencijalima između njenu citoplazmu i spoljašnju sredinu, odnosno sa obe strane površinskih ćelijskih membrana. Potonji je tako polariziran - njegova unutrašnja površina je negativno nabijena u odnosu na vanjsku. Ova razlika potencijala naziva se membranski potencijal. Razlog za ovu potencijalnu razliku je nejednakost koncentracije jona unutar ćelije – u njenoj citoplazmi i izvan ćelije – u okolnoj tkivnoj tečnosti: citoplazma sadrži više jona kalijuma i manje jona natrijuma u odnosu na tkivnu tečnost. U mirovanju, ćelijska membrana je blago propusna za jone Na°. Pri uzbuđenju povećava se propusnost membrane i ona propušta pozitivno nabijene natrijeve ione u ćeliju, što dovodi do smanjenja razlike potencijala membrane (depolarizacija membrane) pa čak i do pojave razlike potencijala suprotnog predznaka. Promjena razlike električnog potencijala tijekom pobude naziva se akcijski potencijal. Generirana električna struja
kada je pobuđena površina tkiva povezana sa neuzbuđenom, to se naziva struja akcije. Ekscitacija je, takoreći, eksplozivni proces koji nastaje zbog promjene propusnosti membrane pod utjecajem iritansa. Ova promjena je u početku relativno mala i praćena je samo blagom depolarizacijom, blagim smanjenjem membranskog potencijala na mjestu primjene stimulacije i ne širi se duž ekscitabilnog tkiva (to je tzv. lokalna ekscitacija). Kada se dosegne kritični - prag - nivo, promjena potencijalne razlike raste poput lavine i brzo - u živcu za nekoliko desethiljaditih dijelova sekunde - dostiže svoj maksimum. Obnavljanje početne razlike potencijala - repolarizacija membrane - nastaje u početku zbog oslobađanja jona kalija iz ćelije. Tada se, zahvaljujući posebnom fiziološkom mehanizmu, takozvanoj natrijum-kalijum pumpi, obnavlja nejednakost koncentracija jona između citoplazme i okoline koja okružuje ćeliju (joni kalija ulaze u ćeliju nazad, a natrijevi je izlaze). Ovaj proces oporavka zahtijeva određeni utrošak energije, koji se nabavlja putem metaboličkih procesa. Karakteristična karakteristika ćelije u trenutku ekscitacije - u periodu maksimalne depolarizacije membrane - je njena nesposobnost da odgovori na novu iritaciju. Stanje neekscitabilnosti ćelije tokom njene ekscitacije naziva se refraktornost. Ekscitacija je valovit proces. Nastala u jednoj ćeliji ili u jednom njenom dijelu, na primjer, u jednom dijelu nervnog vlakna, ekscitacija se širi, prelazi na druge ćelije ili na druge dijelove iste ćelije. Provođenje ekscitacije nastaje zbog činjenice da akcioni potencijal koji je nastao u jednoj ćeliji ili u jednom od njenih područja postaje iritant koji izaziva ekscitaciju željenih područja. Ekscitacija s jedne živčane ćelije na drugu, ili s nervnog vlakna na mišićnu ili žljezdanu ćeliju, se prenosi kemijski. U nervnom završetku nastaju hemijska jedinjenja - prenosioci nervnih impulsa (acetilholin, norepinefrin itd.), koji izazivaju ekscitaciju u ekscitabilnoj ćeliji na kojoj se nalazi nervni završetak. Hemijski prenosioci nervnog impulsa nazivaju se neurotransmiteri.
Fiziologija je usko povezana sa mnogim drugim naukama, bazirajući se u svojim traganjima na podacima nekih nauka, i biće. zauzvrat, osnova za razvoj drugih. Fizika i hemija, biofizika i biohemija, anatomija, histologija i embriologija, opća biologija, genetika i kibernetika - ovo nije potpuna lista disciplina čije je poznavanje od velike važnosti za proučavanje i razumijevanje fizioloških procesa koji se odvijaju u tijelu.
Međutim, vi se spremate da postanete doktori, spremate se da liječite bolesne ljude. A fiziologija proučava funkcije normalnog. zdravo tijelo - zato se i zove "normalna fiziologija". Zašto doktor treba da poznaje funkcije zdravog organizma, ako se u svom radu susreće samo sa bolesnim ljudima? Možda bi bila dovoljna samo "patološka fiziologija" koja se izučava na 3. godini? Mnogi studenti tako misle, a na kraju se ispostavi da nemaju solidno poznavanje ni normalne ni patološke fiziologije, ni kliničkih disciplina.
Mislim da i sami imate ideju zašto doktor treba da poznaje zakone i mehanizme zdravog čoveka. Da li biste popravili televizor ako ne znate principe njegovog rada, ako ne znate kako bi servisni uređaj trebao funkcionirati? Hoćete li sastaviti automobil ako ne znate zakone interakcije svih njegovih dijelova i mehanizama? A čovjek je biološka mašina ogromne složenosti, a njeno popravljanje je veoma težak, delikatan i visokostručan posao! Shvatite disfunkcije koje se javljaju u tijelu tokom razne bolesti, zacrtati ispravne načine njihovog liječenja moguće je samo uz dobro poznavanje fizioloških procesa koji se odvijaju u normalnom, zdravom tijelu.
Značaj fiziologije nije ograničen samo na proučavanje normalnih fizioloških parametara organizma. Samo to bi pomoglo doktoru samo da utvrdi da li osoba ima odstupanja od norme. Ali fiziologija oprema doktora mehanizmima za ispravljanje poremećenih funkcija, omogućavajući takozvani "patogenetski" tretman, koji eliminiše uzrok razvoja bolesti, a ne njene simptome. Savremeni lekar ne može da prepiše lečenje po principu "iz glave, iz stomaka, od hladnoće". U procesu proučavanja pacijenta na osnovu poznavanja fiziologije, on mora pronaći uzrok bolesti i, opet oslanjajući se na fiziologiju, izvršiti takvu intervenciju, koja, i samo koja, dovodi do obaveznog oporavka.
Poznavanje fizioloških metoda proučavanja funkcija omogućava doktoru da odabere one koje su trenutno potrebne, pomaže da se pravilno procijene rezultati ovih studija i iz velikog arsenala lijekovi i medicinske metode za odabir najefikasnijih u ovoj situaciji.
Proučavanje ljudskih bolesti doprinosi razumijevanju mnogih normalnih fizioloških procesa. I.P. Pavlov je o tome pisao: "Svijet patoloških pojava je beskonačan niz svih mogućih, posebnih, ... kombinacija fizioloških pojava koje se inače ne dešavaju. To je kao niz fizioloških eksperimenata prirode i života, ovaj je često takva kombinacija fenomena koja ne bi pala na pamet modernim fiziolozima, a koja se ponekad nije mogla ni namjerno reproducirati tehnička sredstva moderna fiziologija.Klinika postavlja nove zadatke za fiziologiju i istovremeno je bogat izvor novih fizioloških činova.Značaj fiziologije za medicinu i medicine za fiziologiju je toliki da samo njihov bliski spoj može podići kompleksno zdanje nauke o ljudsko tijelo.
Savremena fiziologija je eksperimentalna nauka: fiziološko istraživanje se zasniva na iskustvu, eksperimentu. U eksperimentu se vrši veštačko mešanje u određene procese u organizmu kako bi se razjasnila uloga, značaj i međusobna povezanost pojedinih organa i sistema. Tako, na primjer, kada je cervikalni simpatički nerv iritiran električnom strujom, suzile su žile uha na operiranoj strani, a transekcija ovog živca uzrokuje naglo proširenje ovih žila. Ovi eksperimenti omogućili su fiziologu Claudeu Bernardu da po prvi put uspostavi nervnu regulaciju lumena krvnih sudova.
U arsenalu fiziologije postoje metode kao što su ekstirpacija, transplantacija, denervacija, metoda primjene ligatura i anastomoza, tehnike fistule, metode iritacije i transekcije, perfuzija i registracija funkcija itd. Većina ovih tehnika zahtijeva autopsiju životinje ili operaciju. Koriste se u akutnim ili kroničnim eksperimentima.
Arsenal istraživačkih metoda koji se koristi u eksperimentima na životinjama ne može se koristiti u istraživanju ljudsko tijelo. Stoga su informacije o funkcijama mnogih organa kod ljudi donedavno bile uglavnom ograničene na podatke dobivene u eksperimentima na životinjama. Trenutno se situacija na mnogo načina promijenila. Dostignuća moderne fizike, radiotehnike, elektronike i kibernetike dala su fiziolozima u ruke takve uređaje koji omogućavaju, bez uplitanja u ljudsko tijelo, snimanje rada njegovih organa. Mnoge od ovih uređaja ćete vidjeti tokom studija na Odsjeku za normalnu fiziologiju.
Opći obrasci regulacije funkcija.
Predmet fiziologije su prvenstveno funkcije tijela i njegovih dijelova. Stoga, okrećući se izlaganju fiziologije, moramo se prije svega zadržati na pojmovima ORGANIZMA i FUNKCIJE.
ORGANIZAM je samostalna jedinica organskog svijeta, koja je samoregulirajući sistem koji reagira na različite promjene u vanjskoj sredini u cjelini, sposoban da postoji samo uz stalnu interakciju s vanjskim okruženjem i kao rezultat samoobnavljanja. takve interakcije.
Fiziološke FUNKCIJE su manifestacije vitalne aktivnosti koje imaju adaptivnu vrijednost i usmjerene su na postizanje određenog rezultata korisnog za tijelo. Obavljajući različite funkcije, tijelo se prilagođava vanjskom okruženju ili prilagođava okolinu svojim potrebama.
Glavna funkcija živog organizma je metabolizam i energija (metabolizam). Ovaj proces se sastoji od kombinacije hemijskih i fizičke promjene, u transformaciji supstanci i energije, koja se stalno i kontinuirano odvija u tijelu i svim njegovim strukturama. Metabolizam je neophodan uslov za život. On je taj koji razlikuje živo od neživog. Život je moguć samo dok postoji metabolizam koji održava postojanje žive protoplazme i njeno samoobnavljanje. Prestanak metabolizma dovodi do uništenja protoplazme i smrti organizma.
Sve ostale fiziološke funkcije povezane su s metabolizmom, bilo da se radi o rastu, razvoju, reprodukciji, ishrani, probavi, disanju, izlučivanju, kretanju ili reakcijama na promjene u vanjskom okruženju. Njihova aktivnost je prvenstveno usmjerena na održavanje optimalnih metaboličkih uslova. (Osiguranje normalnog rada tzv. metaboličkog kotla). Istovremeno, osnova bilo koje funkcije je određeni skup transformacija materije i energije. Ovo se podjednako odnosi i na funkcije jedne ćelije, tkiva, organa ili organizma u celini.
Svakom organizmu, jednoćelijskom ili višećelijskom, potrebni su određeni uslovi postojanja koje mu obezbeđuje ono stanište (tzv. spoljašnja sredina) na koje se ova vrsta živih bića prilagođavala tokom čitavog puta svog evolucionog razvoja. Funkcije tijela se normalno obezbjeđuju samo ako mu vanjsko okruženje daje mogućnost da prima hranu, na određenoj temperaturi, barometarskom pritisku, intenzitetu i spektru svjetlosti itd.
Pritom treba imati na umu da su granice okolišnih fluktuacija koje toleriše organizam viših životinja mnogo šire od onih koje su potrebne za normalno funkcioniranje većine njegovih stanica. Razlog tome je što je stanište ćelija tela njegovo unutrašnje okruženje, koje se mnogo manje menja od spoljašnjeg. Unutrašnja sredina tijela je krv, limfa i tkivna tekućina u kojoj žive stanice.
Funkcije tjelesnih stanica su normalne samo uz relativnu konstantnost
osmotski pritisak, sastav elektrolita, određena koncentracija vodikovih jona, nutritivni i energetski resursi. Konstantnost hemijskog sastava i fizičko-hemijskih svojstava unutrašnje sredine je važna karakteristika viših životinjskih organizama. Da bi označio ovu postojanost, Cannon je predložio termin homeostaza. Izraz homeostaze je prisustvo niza bioloških konstante, tj. stabilni kvantitativni pokazatelji koji karakteriziraju normalno stanje tijela. U zavisnosti od njihove vrednosti, razlikuju se tvrde i meke (plastične) konstante. Uočavajući postojanost sastava, fizičko-hemijskih i bioloških svojstava unutrašnje sredine, treba naglasiti da ona nije apsolutna, već relativna i dinamična. Ova konstantnost se postiže neprekidnim radom niza organa i sistema, usled čega dolazi do pomaka u sastavu i fizičko-hemijskim svojstvima unutrašnje sredine do kojih dolazi pod uticajem promena u spoljašnjoj sredini i vitalne aktivnosti organizma. organizam je usklađen.
Homeostaza ima određene granice. Tokom posebno dugog boravka, u uslovima značajno drugačijim od onih na koje je organizam prilagođen, dolazi do poremećaja homeostaze i do pomaka nespojivih sa životom. Čak i male povrede homeostaze dovode do patologije, stoga je određivanje relativno konstantnih fizioloških konstanti (pH, krvni pritisak, respiratorna frekvencija, srčana frekvencija, MOD, itd.) od velike dijagnostičke vrijednosti. Stoga se rad na održavanju homeostaze odvija stalno i stalno reguliran od strane relevantnih regulatornih sistema, o čemu ćemo kasnije govoriti.
Uloga različitih organa i njihovih sistema u održavanju homeostaze je različita. O njima ćemo također govoriti u relevantnim dijelovima kursa.
Kao što je već spomenuto, karakteristična karakteristika svakog živog organizma je da je to samoregulirajući sistem koji u cjelini reagira na različite utjecaje. Princip samoregulacije je da je odstupanje bilo koje konstante od normalnog nivoa samo po sebi signal za ispravljanje ovih pomaka. Samoregulacija se postiže interakcijom svih ćelija tijela, njegovih tkiva i organa. Ova interakcija organa posebno je jasno izražena u radu tzv. funkcionalni sistemi. Takav sistem čine organi čija zajednička aktivnost osigurava prilagođavanje određenim uvjetima okoline, osiguravajući zadovoljenje bilo koje unutrašnje potrebe.
Ispod unutrašnje potrebe od sada ćemo razumeti svako manje ili više dugotrajno odstupanje jedne ili druge konstante njegovog unutrašnjeg okruženja od nivoa koji obezbeđuje njegovu normalnu životnu aktivnost. Upravo su biološke potrebe prvi poticaj u lancu procesa samoregulacije različitih tjelesnih funkcija.
Potrebe živog organizma mogu biti bezbrojne. Međutim, svi se oni spajaju u velike grupe - biološke, socijalne, seksualne, prehrambene, odbrambene itd. Zadovoljenje jedne ili druge potrebe i predstavlja za svaki živi organizam određenu koristan rezultat njegova adaptivna aktivnost, tj. funkcije.
Regulacija je usmjerena promjena u funkcijama različitih organa i tkiva. Sve funkcije u tijelu reguliraju dva glavna regulatorna mehanizma - nervozna i humoralna.
humoralni mehanizam Zasniva se na činjenici da se u različitim ćelijama i organima, tokom procesa metabolizma, stvaraju hemikalije različite prirode i fiziološkog dejstva. Ulazeći u tkivnu tečnost, a zatim u krv, raznosi se po celom telu i može uticati na ćelije tkiva. Poseban slučaj humoralne regulacije je endokrina, koju provode endokrine žlijezde.
neuronski mehanizam Regulacija leži u činjenici da se startne ili modulirajuće komande šalju preko nerava do svih ćelija i organa, mijenjajući njihovu aktivnost u smjeru potrebnom za tijelo. Promjene stanja nekih ćelija i organa kroz nervni sistem na refleksni način izazivaju promjene u funkcijama drugih organa. Ovaj mehanizam regulacije je savršeniji, jer se interakcija ćelija kroz nervni sistem odvija mnogo brže od humoralno-hemijske, a osim toga, nervni impulsi uvijek imaju na umu određenog primatelja.
Dakle, dva opšti princip regulacija svih funkcija u tijelu:
1. Sve funkcije u tijelu reguliše nervni i/ili humoralni sistem.
2. Regulisanje funkcija vrši se po principu samoregulacije.
Oba ova principa se najjasnije i najpotpunije manifestuju u aktivnostima tzv funkcionalni sistemi(FS), koji se konstantno formiraju kada se u organizmu pojavi potreba i osiguravaju njeno optimalno zadovoljenje.
Funkcionalni sistem je skup heterogenih organa i tkiva koji su ujedinjeni na funkcionalnoj osnovi i daju, u interakciji, kvalitativno nove funkcije i oblike aktivnosti, s rezultatom koji je svojstven sistemu kao cjelini, a ne svojstven njegovim dijelovima odvojeno. FS je dinamična, samoregulirajuća organizacija, aktivnosti svih sastavni elementišto doprinosi postizanju adaptivnog rezultata vitalnog za tijelo.
Sastav FS može uključivati različite organe i tkiva čija aktivnost može dovesti do obnove poremećene homeostaze. Funkcionalni sistemi su organizovani ne prema anatomskim, već prema fiziološkim karakteristikama. Glavni faktor koji formira sistem je cilj, rezultat budućih aktivnosti FS. Kako napredujemo kroz kurs fiziologije, karakteriziraćemo različite posebne FS (održavanje pH, osmotski pritisak, koncentracija nutrijenata u krvnom tlaku, itd.) Sada moramo razmotriti opću shemu strukture bilo kojeg FS. ovu opštu shemu, specifične regulatorne i izvršne mehanizme i rezultate njihovih aktivnosti, da izgrade šemu bilo koje FS.
Centralni sistemotvorni faktor svakog PS je rezultat njegovog delovanja, koji određuje normalne uslove za tok metaboličkih procesa u telu kao celini. Iz ovoga proizilazi da je upravo rezultat svojevrsna "vizit karta" svakog FS-a.
U živom organizmu mogu se razlikovati 3 grupe korisnih adaptivnih rezultata.
Prvu grupu čine unutrašnje konstante organizma, homeostatski indikatori koji određuju njegovu normalnu vitalnu aktivnost: pH, koncentracija soli, nutrijenata, gasova itd.
Drugu grupu čine rezultati adaptivne aktivnosti organizma u životnoj sredini, u cilju zadovoljavanja njegovih unutrašnjih bioloških potreba, očuvanja vrste i roda (svrsishodno ponašanje koje dovodi do gašenja žeđi, gladi i sl.).
Treća grupa su rezultati društvene aktivnosti osobe usmjerene na zadovoljavanje njegovih društvenih potreba.
U zavisnosti od svojstava rezultata, FS će biti jednostavniji ili složeniji, ali njegova opšta šema ostaje ista (i) humoralna povratna sprega (aferentacija) selektivno mobiliše posebne regulatorne aparate. Potonji, preko izvršnih uređaja, ponovo vraćaju korisni adaptivni rezultat na traženi nivo. Svi ovi procesi se odvijaju kontinuirano uz stalno informiranje centra o uspješnosti postizanja korisnog adaptivnog rezultata. Isti aktuatori i periferni organi mogu se mobilizirati za obavljanje različitih tjelesnih funkcija i biti dio različitih FS.
Živi organizam je najsloženiji mehanizam, koji se sastoji od različitih FS koji su blisko uklopljeni u svoj rad i imaju zajedničke tačke kontakt i određena hijerarhija. Međutim, u svakom konkretnom slučaju, u svakom konkretnom trenutku, uvijek postoji dominantna FS, koja određuje aktivnost organizma u datom trenutku i podređuje aktivnost ostalih funkcionalnih sistema.
Rad mnogih FS tijela objedinjuje krv, kao tačka koncentracije konačnih rezultata mnogih FS. Kako je krv unutrašnja sredina tijela, budući da su njene konstante homeostatske i u velikoj mjeri rigidne, proučavanje predmeta fiziologija počinjemo odlomkom „Fiziologija krvnog sistema“.
PREDAVANJAU LJUDSKOJ FIZIOLOGIJI
Fiziologija kao nauka. Predmet, zadaci, metode, istorija fiziologije
Fiziologija (physis - priroda) je nauka o normalnim životnim procesima organizma, njegovim sastavnim fiziološkim sistemima, pojedinim organima, tkivima, ćelijama i subcelularnim strukturama, mehanizmima regulacije ovih procesa i uticaju prirodnih faktora sredine na funkcije. tijela.
Na osnovu toga, općenito, predmet fiziologije je zdrav organizam. Zadaci fiziologije uključeni su u njenu definiciju. Glavna metoda fiziologije je eksperiment na životinjama. Postoje 2 glavne vrste eksperimenata ili eksperimenata:
1. Akutno iskustvo ili vivisekcija (živo rezanje). U procesu se izvodi hirurška intervencija, ispituju se funkcije otvorenog ili izoliranog organa. Nakon toga ne traže opstanak životinje. Trajanje akutnog eksperimenta je od nekoliko desetina minuta do nekoliko sati (primjer).
2. Hronična iskustva. U toku kroničnih eksperimenata vrši se hirurška intervencija kako bi se dobio pristup organu. Tada postižu zacjeljivanje kirurških rana i tek nakon toga kreću u istraživanja. Trajanje kroničnih eksperimenata može biti mnogo godina (primjer).
Ponekad se razlikuje subakutni eksperiment (primjer).
Istovremeno, medicina zahtijeva informacije o mehanizmima funkcioniranja ljudskog tijela. Stoga, I.P. Pavlov je napisao: "Eksperimentalni podaci mogu se primijeniti na osobu samo s oprezom, stalno provjeravajući činjenično stanje sličnosti s aktivnostima ovih organa kod ljudi i životinja." Shodno tome, bez posebnih zapažanja i eksperimenata na osobi, proučavanje njegove fiziologije Zbog toga se izdvaja posebna fiziološka nauka - fiziologija čoveka, ljudska fiziologija ima predmet, zadatke, metode i istoriju. Predmet fiziologije čoveka je zdravo ljudsko telo.
Njeni zadaci:
1. Proučavanje mehanizama funkcionisanja ćelija, tkiva, organa, sistema ljudskog tela u celini
2. Proučavanje mehanizama regulacije funkcija organa i sistema tijela.
3. Otkrivanje reakcija ljudskog tijela i njegovih sistema na promjene u vanjskom i unutrašnjem okruženju.
Budući da je fiziologija u cjelini eksperimentalna nauka, glavni metod ljudske fiziologije je također eksperiment. Međutim, eksperimenti na ljudima se bitno razlikuju od eksperimenata na životinjama. Prvo, velika većina istraživanja na ljudima radi se neinvazivnim metodama, tj. bez intervencija u organima i tkivima (npr. EKG, EEG, EMG, krvni testovi itd.). Drugo, eksperimenti na ljudima se izvode samo kada ne štete zdravlju i uz pristanak ispitanika. Ponekad se akutni eksperimenti izvode na osobi u klinici kada to zahtijevaju dijagnostički zadaci (primjer). Međutim, treba napomenuti da bi bez podataka klasične fiziologije nastanak i razvoj ljudske fiziologije bio nemoguć (spomenici žabi i psu). Više I.P. Pavlov je, ocjenjujući ulogu fiziologije za medicinu, napisao: "Shvaćeno u grubom smislu riječi, fiziologija i medicina su neodvojive, poznavanje fiziologije je neophodno za doktora bilo koje specijalnosti." I još da će „Medicina, koja se iz dana u dan neprestano obogaćuje, novim fiziološkim činjenicama, konačno postati ono što bi idealno trebalo da bude, odnosno sposobnost da se popravi pokvareni ljudski mehanizam i primeni znanja iz fiziologije“ (primeri sa klinike) . Još jedan poznati ruski fiziolog prof. V.Ya. Danilevsky je napomenuo: „Što se preciznije i potpunije utvrde znakovi norme za tjelesni i mentalni život osobe, to će liječnikova dijagnoza biti ispravnija za njegove patološke abnormalnosti.
Fiziologija, kao fundamentalna biološka nauka, usko je povezana sa drugim fundamentalnim i biološke nauke. Konkretno, bez poznavanja zakona fizike nemoguće je objasniti bioelektrične fenomene, mehanizme percepcije svjetlosti i zvuka. Bez upotrebe hemijskih podataka nemoguće je opisati procese metabolizma, probave, disanja itd. Stoga se na granici ovih nauka sa fiziologijom izdvajaju kćerke nauke biofizika i biohemija.
Budući da su struktura i funkcija neodvojive, a funkcija je ta koja određuje formiranje strukture, fiziologija je usko povezana sa morfološkim naukama: citologijom, histologijom, anatomijom.
Kao rezultat proučavanja djelovanja različitih kemikalija na tijelo, farmakologija i toksikologija su se iz fiziologije izrodile u samostalne nauke. Akumulacija podataka o kršenju mehanizama funkcioniranja tijela u različitim bolestima poslužila je kao osnova za nastanak patološke fiziologije.
Razlikovati opštu i posebnu fiziologiju. Opća fiziologija proučava osnovne obrasce života tijela, mehanizme osnovnih procesa kao što su metabolizam i energija, reprodukcija, procesi ekscitacije itd. Posebna fiziologija proučava funkcije specifičnih ćelija, tkiva, organa i fizioloških sistema. Stoga ističe dijelove kao što su fiziologija mišićnog tkiva, srca, bubrega, probave, disanja itd. Osim toga, u fiziologiji postoje sekcije koje imaju specifičan predmet proučavanja ili posebne pristupe proučavanju funkcija. To uključuje evolucijsku fiziologiju (objašnjenje), komparativnu fiziologiju, fiziologiju starosti.
Postoji niz primijenjenih grana u fiziologiji. Ovo je, na primjer, fiziologija domaćih životinja. U ljudskoj fiziologiji razlikuju se sljedeći primijenjeni dijelovi:
1. Fiziologija starosti. Proučava starosne karakteristike tjelesnih funkcija.
2. Fiziologija porođaja.
3. Klinička fiziologija. Ovo je nauka koja koristi fiziološke metode i pristupe za dijagnostiku i analizu patoloških abnormalnosti.
4. Vazduhoplovna i svemirska fiziologija.
5. Fiziologija sporta.
Ljudska fiziologija je usko povezana sa kliničkim disciplinama kao što su terapija, hirurgija, akušerstvo, endokrinologija, psihijatrija, oftalmologija itd. Na primjer, ove nauke koriste brojne metode koje su razvili fiziolozi za dijagnostiku. Odstupanja normalnih parametara tijela osnova su za otkrivanje patologije.
Neki dijelovi ljudske fiziologije su osnova psihologije. Ovo je fiziologija centralnog nervnog sistema, viša nervna aktivnost, senzorni sistemi, psihofiziologija.
Istorija fiziologije je detaljno opisana u udžbeniku, ur. Tkachenko
^ MEHANIZMI REGULACIJE TJELESNIH FUNKCIJA
Humoralna i nervna regulacija. Reflex. Refleksni luk. Osnovni principi teorije refleksa
Sve tjelesne funkcije reguliraju dva sistema regulacije: humoralni i nervni. Filogenetski starija humoralna regulacija je regulacija pomoću fiziološki aktivnih supstanci (PAS) koje kruže u tjelesnim tekućinama - krvi, limfi, međućelijskoj tekućini. Faktori humoralne regulacije su:
1. Neorganski metaboliti i joni. Na primjer, kationi kalcija, vodika, ugljičnog dioksida.
2. Hormoni endokrinih žlijezda. Proizvode ga specijalizovane endokrine žlezde. To su insulin, tiroksin itd.
3. Lokalni ili tkivni hormoni. Ove hormone proizvode posebne ćelije zvane parakrine, transportuju se tkivnom tečnošću i deluju samo na maloj udaljenosti od ćelija koje luče. To uključuje supstance kao što su histamin, serotonin, gastrointestinalni hormoni i druge.
4. Biološki aktivne supstance koje obezbeđuju kreativne veze između ćelija tkiva. To su proteinske makromolekule koje luče. Oni reguliraju diferencijaciju, rast i razvoj svih stanica koje čine tkivo i osiguravaju funkcionalnu povezanost stanica u tkivu. Takvi proteini su, na primjer, keyoni, koji inhibiraju sintezu DNK i diobu stanica.
Glavne karakteristike humoralne regulacije:
1.Mala brzina regulatornog dejstva povezana sa malom brzinom strujanja odgovarajućih telesnih tečnosti.
2. Sporo povećanje jačine humoralnog signala i sporo smanjenje. To je zbog postepenog povećanja koncentracije PAS-a i njihovog postepenog uništavanja.
3. Odsustvo specifičnog tkiva ili ciljnog organa za djelovanje humoralnih faktora. Djeluju na sva tkiva i organe duž toka tekućine, u čijim ćelijama se nalaze odgovarajući receptori.
Nervna regulacija je regulacija tjelesnih funkcija putem refleksa koje provodi nervni sistem.
Koncept refleksnog principa aktivnosti nervnog sistema prvi je razvio u 17. veku francuski prirodnjak Rene Descartes. Predložio je hipotetičku shemu za formiranje nevoljnog pokreta (mehaničko predstavljanje). Termin "refleks" (reflektivno djelovanje) uveo je u fiziologiju 1771. godine Unzer. J. Prohaska je 1800. godine razvio dijagram najjednostavnijeg refleksnog luka. NJIH. Sechenov je proširio koncept "refleksa" na bilo koju, uključujući višu živčanu aktivnost (HNA). Istovremeno je polazio od 2 odredbe: 1. svaka aktivnost organizma na kraju se svodi na kretanje. 2. Svi pokreti su po poreklu refleksi. I.P. Pavlov je eksperimentalno potkrijepio gledište o refleksu kao glavnom činu svake nervne aktivnosti. On je također podijelio sve reflekse, prema mehanizmu formiranja, na bezuvjetne i uslovne. Glavne karakteristike teorije refleksa I.P. Pavlov je formulisao u svom djelu "Odgovor fiziologa psiholozima." Ono uključuje tri osnovna principa:
1. Princip determinizma. Piše "nema akcije bez razloga". One. svaki refleksni čin je posljedica djelovanja stimulusa na organizam.
2. Princip analize i sinteze. Analiza se stalno odvija u mozgu; diskriminacija signala, kao i sinteza, tj. njihova interakcija i holistička percepcija.
3. Princip strukture. AT nervni sistem ne postoje procesi koji nemaju određenu strukturnu lokalizaciju.
Morfološka osnova svaki refleks je refleksni luk ili refleksni put. Refleksni luk (RD) je put refleksne reakcije, tj. nervne signale. Refleksni luk somatskog (motornog) refleksa sastoji se od sljedećih glavnih karika:
1. Receptor koji percipira iritaciju
2. Aferentno ili uzlazno ili senzorno nervno vlakno
3. Nervni centar u C.N.S.
4. Eferentno ili silazno, motorno nervno vlakno
5. Izvršni organ "efektor"
U nizu refleksnih lukova nalazi se neuron povratne sprege (6), odnosno neuron reverzne aferentacije, koji reagira na refleksni odgovor i kontrolira ga.
U somatskom refleksnom luku mogu se razlikovati neuroni koji obavljaju određene funkcije. Konkretno, u najjednostavnijem monosinaptičkom refleksu postoje samo 2 neurona: osjetljivi i motor. U najjednostavnijem polisinaptičkom refleksnom luku koji razmatramo postoje: a) osjetljivi neuron, b) interkalarni neuron, c) izvršni neuron. Rice. Postoje stotine i hiljade neurona u kompleksnim polisinaptičkim refleksnim lukovima.
U luku autonomnog refleksa postoje sljedeće veze:
1. Receptor
2. Aferentno nervno vlakno.
3. Nervni centar (na primjer) za simpatičke reflekse u bočnim rogovima kičmena moždina)
4. Preganglijska nervna vlakna
5. Autonomna ganglija
6.Postganglijska nervna vlakna
7. Izvršni organ. Rice.
Biološki i funkcionalni sistemi
Tokom 1950-ih i 1960-ih godina, kanadski biolog Ludwig Bertalanffy je, koristeći matematičke i kibernetičke pristupe, razvio osnovne principe za rad bioloških sistema. To uključuje:
1. Integritet, tj. nesvodljivost svojstava sistema na prost zbir svojstava njegovih delova. One. nemoguće je opisati svojstva biološkog sistema kroz funkcije njegovih pojedinačnih elemenata (primjer).
2. Strukturni. Mogućnost objašnjenja funkcija sistema kroz njegovu strukturu (primjer).
3. Hijerarhija, podređenost elemenata sistema jedni drugima od vrha do dna. One. gornje komponente sistema upravljaju osnovnim (primjer).
4. Odnos između sistema i okoline (primjer).
Međutim, Bertalanffy nije otkrio najvažnije – faktor koji formira sistem. Stoga, glavna uloga u identifikaciji sistemskih obrazaca svojstvenih živim organizmima pripada akademiku P.K. Anokhin. U fiziologiji je koncept fizioloških sistema dugo postojao. Ovo je kompleks morfološki i funkcionalno ujedinjenih organa koji imaju zajedničke regulatorne mehanizme i obavljaju homogene funkcije (primjer). Međutim, P.K. Anokhin je ustanovio da postoje i drugi sistemi u tijelu, na primjer, koji pružaju vitalnu podršku. važnih telesnih parametara. On ih je nazvao funkcionalnim sistemima (FUS). Prema P.K. Anokhin FUS je skup organa i tkiva koji osiguravaju postizanje cilja u određenoj vrsti životne aktivnosti. Ovaj cilj se zove korisni adaptivni rezultat (PPR). To može biti bilo koji parametar unutrašnjeg okruženja, na primjer, tjelesna temperatura, normalan sadržaj kisika u krvi, itd., rezultat ponašanja koje zadovoljava biološku, na primjer, nutritivnu potrebu, rezultat društvene aktivnosti osobe. Važno je da ljekar razumije FUS koji obezbjeđuje homeostazu.
Upravo je PPR faktor koji objedinjuje različite organe i sisteme tijela u jedinstvenu cjelinu - FUS. Povezivanje organa u FUS se ne odvija prema morfološkoj, već prema funkcionalnoj osobini. Stoga, FUS može uključivati organe i tkiva iz različitih fizioloških sistema. Štaviše, isti organi mogu biti uključeni u nekoliko FUS odjednom. Osim toga, za razliku od fizioloških sistema, FUS može biti i naslijeđen i formiran tokom individualni život. Opća FUS shema za održavanje parametara homeostaze uključuje sljedeće elemente:
2. PPR receptori
3. Aferentni put
4.Nervni centar
5. Vegetativna regulacija
6. Humoralna regulacija
7. Regulacija ponašanja
8. Metabolizam (sl.)
Ako, pod utjecajem bilo kojeg uzroka, PPR prelazi granice fiziološke norme, PPR receptori se pobuđuju. Nervni impulsi iz njih dolaze do nervnog centra koji regulira ovu funkciju. Odatle idu u izvršne organe koji osiguravaju održavanje odgovarajućeg parametra homeostaze. Istovremeno se pokreću humoralni mehanizmi regulacije. Kada, uprkos tome, PPR ne dosegne početni nivo, nervni impulsi iz nervnog centra počinju da teče u moždanu koru. Kao rezultat ekscitacije njegovih neurona, uključuje se vanjska karika samoregulacije tijela, tj. regulacija ponašanja. Ovo je svrsishodna promjena u ponašanju živog bića. Kao rezultat ovih regulatornih radnji, PPR dolazi na početni nivo, tj. fiziološka norma. PPR je direktno pod utjecajem metabolizma. S druge strane, sam PPR ima direktan utjecaj na metabolizam procesi. Primjeri funkcioniranja raznih FUS-a.
^ Principi samoregulacije organizma. Koncept homeostaze
i homeokineza
Sposobnost samoregulacije je glavno svojstvo živih sistema.Neophodno je stvoriti optimalne uslove za interakciju svih elemenata koji čine tijelo, kako bi se osigurao njegov integritet. Postoje četiri glavna principa samoregulacije:
1. Princip neravnoteže ili gradijenta. Biološka suština života leži u sposobnosti živih organizama da održavaju dinamičko neravnotežno stanje u odnosu na okolinu. Na primjer, tjelesna temperatura toplokrvnih životinja je viša ili niža od okoline. U ćeliji je više kalijevih katjona, a van nje natrijuma itd. Održavanje potrebnog nivoa asimetrije u odnosu na okolinu osigurava se procesima regulacije.
2. Princip zatvorene upravljačke petlje. Svaki živi sistem ne samo da reaguje na stimulus, već i ocenjuje korespondenciju odgovora na trenutni stimulus. One. što je iritacija jača, to je veća reakcija i obrnuto. Ova samoregulacija se provodi zbog obrnutih pozitivnih i negativnih povratne informacije u nervnom i humoralnom sistemu regulacije. One. regulacijski krug je zatvoren u prsten. Primjer takve veze je stražnji aferentni neuron u lukovima motornih refleksa.
3. Princip predviđanja. Biološki sistemi su u stanju da predvide ishode odgovora na osnovu prethodnog iskustva. Primjer je izbjegavanje bolnih podražaja nakon prethodnih.
4. Princip integriteta. Za normalno funkcionisanje živog sistema potreban je njegov strukturni integritet.
Doktrinu homeostaze razvio je C. Bernard. Godine 1878. iznio je hipotezu o relativnoj postojanosti unutrašnjeg okruženja živih organizama. 1929. W. Cannon je pokazao da je sposobnost tijela da održi homeostazu rezultat regulatornih sistema u tijelu. Takođe je skovao termin homeostaza. Konstantnost unutrašnje sredine organizma (krv, limfa, tkivna tečnost, citoplazma) i stabilnost fizioloških funkcija rezultat je homeostatskih mehanizama. Kod kršenja homeostaze, kao što je ćelijska, dolazi do degeneracije ili smrti ćelija. Ćelijska, tkivna, organska i drugi oblici homeostaze regulišu se i koordiniraju humoralnom, nervnom regulacijom, kao i nivoom metabolizma.
Parametri homeostaze su dinamični i mijenjaju se u određenim granicama pod utjecajem okolišnih faktora (npr. pH krvi, sadržaj respiratornih plinova i glukoze u njoj itd.). To je zbog činjenice da živi sistemi ne balansiraju samo vanjske utjecaje, već im se aktivno suprotstavljaju. Sposobnost održavanja postojanosti unutrašnjeg okruženja s promjenama u vanjskom glavno je svojstvo koje razlikuje žive organizme od nežive prirode. Stoga su vrlo neovisni o vanjskom okruženju. Što je veća organizacija živog bića, to je ono više nezavisno od spoljašnje sredine (primer).
Kompleks procesa koji obezbeđuju homeostazu naziva se homeokineza. Izvode ga sva tkiva, organi i sistemi tijela. Međutim, funkcionalni sistemi su od najveće važnosti.
^ Dobne karakteristike fizioloških funkcija
i neurohumoralna regulacija
U procesu razvoja organizma nastaju i kvantitativne i kvalitativne promjene. Na primjer, povećava se broj mnogih ćelija i njihove veličine. Istovremeno, kao rezultat komplikacije strukture tijela, pojavljuju se nove funkcije. Na primjer, mozak djeteta u razvoju stječe sposobnost apstraktnog razmišljanja.
Sljedeći fenomeni su u osnovi starosnih promjena u funkcijama tjelesnih sistema:
1. Neravnomjerno ili heterohrono sazrijevanje organa i sistema tijela. 2 . Inscenirani skokovi u godinama.
3. Ubrzanje. One. ubrzanje brzine biološkog razvoja u određenom periodu.
Sazrevanje pojedinih organa i sistema se ne odvija istovremeno (heterohrono). Kod novorođenčeta se pre svega razvijaju oni fiziološki i funkcionalni sistemi koji obezbeđuju opstanak organizma tokom perioda. prelazak iz intrauterinog u vanmaternično postojanje. Na osnovu zapažanja o formiranju funkcionalnih sistema u procesu ontogeneze, akademik P.K. Anohin je stvorio doktrinu sistemogeneze. Heterohronost razvoja organa i sistema može se ilustrovati na primjeru motoričkog aparata djeteta. U početku se formiraju refleksi koji omogućavaju držanje glave, zatim sposobnost sjedenja, zatim stajanja i na kraju hodanja. Program individualni razvoj izvodi genetski aparat. U određenim starosnim fazama dolazi do izražaja, tj. aktivacija dobro definisanih gena. Kao rezultat, ubrzava se sazrijevanje određenog sistema ili tjelesne funkcije. To se manifestira skokom u dobi ili kritičnim periodom. Na primjer, primjećuje se nagla promjena u strukturi i funkciji organa i sistema tokom puberteta.
Ubrzanje je povezano sa uticajem okoline i društvenih faktora na organizam. Prati ga brzi rast skeleta, mišića, unutrašnje organe, pubertet.
Formiranje i razvoj tijela završava se za oko 20 godina. Ljudi starosti od 20 do 55-60 godina smatraju se zrelim godinama.U tom periodu se u potpunosti formiraju sve tjelesne funkcije, funkcionalna aktivnost organa i sistema je približno na istom nivou. Za starije osobe u dobi od 65-75 godina karakteristična je pojava involutivnih preuređivanja. Jedan od glavnih znakova starenja je smanjenje bazalnog metabolizma, zbog čega su metabolički procesi u stanicama poremećeni. Bazalni metabolizam se smanjuje kao rezultat smanjenja broja mitohondrija u stanicama. Smatra se da je vrijednost bazalnog metabolizma jedan od najvažnijih faktora koji određuju trajanje ljudskog života. Nakon 75 godina nastupa starost. Aktivnost svih fizioloških procesa naglo se smanjuje. Kao rezultat toga, javljaju se mnoge senilne bolesti, poput ateroskleroze.
Mehanizmi neurohumoralne regulacije se također mijenjaju s godinama. Novorođenče ima ograničen broj složenih bezuslovnih refleksa, a nema uslovnih. U isto vrijeme, ćelije su vrlo osjetljive na humoralne faktore. Sa rastom se poboljšava refleksna aktivnost centralnog nervnog sistema. Konkretno, do godine dana formiraju se vrlo složeni refleksi koji pružaju govor. Istovremeno, početna osjetljivost stanica na humoralne faktore se smanjuje. zrela osoba ima visoko organizovane mehanizme neurohumoralne regulacije. U starijoj dobi, brzina i težina refleksnih reakcija se smanjuje. Slabljenje nervnih uticaja na organe i tkiva nastaje usled destruktivnih promena u nervnim završecima i sinapsama u centralnom nervnom sistemu i na periferiji. Istovremeno, zbog promjena u receptorskom aparatu stanica, smanjuje se njihova osjetljivost na niz humoralnih faktora.
Za pedijatrijski fakultet važno je poznavati periode detinjstva. Dodijelite (prema Arshavskyju):
1. Neonatalni period traje 7-8 dana.
2. Period dojenje- 5-6 meseci
3. Period mešovite ishrane od 6 do 12 meseci.
4. Dječji period 1 godina - 3 godine
5. Period predškolskog uzrasta 3 - 7 godina.
6. Period osnovnoškolskog uzrasta 7-12 godina
7. Period starijeg školskog uzrasta 12-17 godina
8, Period mladosti 17-20 godina
FIZIKA I OLOGIJA I B I O FIZIKA I C A V O Z B U D I M X
C L E T O C
^ Koncept razdražljivosti, razdražljivosti i uzbuđenja. Klasifikacija stimulusa
Razdražljivost je sposobnost ćelija, tkiva, tijela u cjelini da pod utjecajem vanjskih ili unutrašnjih faktora okoline pređu iz stanja fiziološkog mirovanja u stanje aktivnosti. Stanje aktivnosti se manifestuje promjenom fizioloških parametara ćelije, tkiva, organizma, na primjer, promjenom metabolizma.
Ekscitabilnost je sposobnost živog tkiva da na iritaciju odgovori aktivnom specifičnom reakcijom – ekscitacijom, tj. generisanje nervnog impulsa, kontrakcije, sekrecije. One. ekscitabilnost karakterizira specijalizirana tkiva - živčana, mišićna, žljezdana, koja se nazivaju ekscitabilna. Ekscitacija je kompleks procesa ekscitabilnog tkiva koji reaguje na djelovanje iritansa, koji se manifestira promjenom membranskog potencijala, metabolizma itd. Ekscitabilna tkiva su provodljiva. Ovo je sposobnost tkiva da sprovodi ekscitaciju. Najveću provodljivost imaju nervi i skeletni mišići.
Iritans je faktor spoljašnje ili unutrašnje sredine koji deluje na živo tkivo.
Proces izlaganja iritansu ćelije, tkiva, organizma naziva se iritacija.
Svi podražaji su podijeljeni u sljedeće grupe:
1.Po prirodi
A) fizički (struja, svjetlost, zvuk, mehanički utjecaji, itd.)
B) hemijske (kiseline, alkalije, hormoni, itd.)
C) fizički i hemijski (osmotski pritisak, parcijalni pritisak gasova, itd.)
D) biološki (hrana za životinju, jedinku suprotnog pola)
D) društveni (reč za osobu).
2. Prema mjestu udara:
A) vanjski (egzogeni)
B) interni (endogeni)
3. Po snazi:
A) podprag (ne izaziva odgovor)
B) prag (podražaj minimalne sile pri kojoj dolazi do ekscitacije)
C) superprag (snaga iznad praga)
4. Po fiziološkoj prirodi:
A) adekvatna (fiziološka za datu ćeliju ili receptor koji joj se prilagodio u procesu evolucije, npr. svjetlo za fotoreceptore oka).
B) neadekvatan
Ako je reakcija na podražaj refleksna, postoje i:
A) bezuslovni refleksni nadražaji
B) uslovni refleks
^ Zakoni iritacije. parametri ekscitabilnosti.
Reakcija ćelija, tkiva na iritans određena je zakonima iritacije
1. Zakon "sve ili ništa": Kod iritacije ćelija pre praga, odgovor tkiva se ne javlja. Na pragu jačine stimulusa razvija se maksimalni odgovor, stoga povećanje jačine iritacije iznad praga nije popraćeno njegovim povećanjem. U skladu sa ovim zakonom, jedno nervno i mišićno vlakno, srčani mišić, reaguje na podražaje.
2. 2. Zakon snage: Što je jačina stimulusa, jači je odgovor. Međutim, težina odgovora raste samo do određenog maksimuma. Holistički skeletni, glatki mišići poštuju zakon sile, budući da se sastoje od brojnih mišićnih ćelija različite ekscitabilnosti.
3. Zakon trajanja sile. Postoji određeni odnos između jačine i trajanja stimulusa. Što je stimulus jači, to je manje vremena potrebno da dođe do odgovora. Odnos između granične sile i potrebnog trajanja stimulacije odražava se na krivulji sila-trajanje. Iz ove krive može se odrediti niz parametara ekscitabilnosti.A) Prag iritacije je minimalna jačina stimulusa pri kojoj dolazi do ekscitacije.
B) Rebaza je minimalna snaga stimulusa koji izaziva ekscitaciju tokom svog delovanja na neograničeno dugo vreme. U praksi, prag i reobaza imaju isto značenje. Što je niži prag iritacije ili manje rebaze, to je veća ekscitabilnost tkiva.
C) Korisno vrijeme - ovo je minimalno vrijeme djelovanja stimulusa snagom od jedne reobaze tokom kojeg dolazi do ekscitacije.
D) Chronaxia - ovo je minimalno vrijeme djelovanja stimulusa sa silom dvije reobaze, neophodno za početak ekscitacije. Ovaj parametar je predložio da izračuna L. Lapik, radi preciznijeg određivanja indikatora vremena na krivulji sila-trajanje. Što kraće korisno vrijeme ili chronaxia, veća je ekscitabilnost i obrnuto.
U kliničkoj praksi, reobaza i kronaksija se određuju metodom chronaxis za proučavanje ekscitabilnosti nervnih stabala.
4. Zakon gradijenta ili akomodacije. Odgovor tkiva na iritaciju zavisi od njegovog gradijenta, tj. što se jačina stimulusa brže povećava u vremenu, brži se javlja odgovor. Pri maloj stopi povećanja jačine stimulusa, povećava se prag iritacije. Stoga, ako se jačina stimulusa povećava vrlo sporo, neće biti uzbuđenja. Ovaj fenomen se naziva akomodacija.
Fiziološka labilnost (pokretljivost) je veća ili manja učestalost reakcija na koje tkivo može odgovoriti na ritmičku stimulaciju. Što se brže obnavlja njegova ekscitabilnost nakon sljedeće iritacije, to je veća njegova labilnost. Definiciju labilnosti predložio je N.E. Vvedensky. Najveća labilnost u nervima, najmanja u srčanom mišiću.
^ Djelovanje jednosmjerne struje na ekscitabilna tkiva
Po prvi put, pravilnosti djelovanja jednosmjerne struje na neuromišićni lijek proučavao je Pfluger u 19. stoljeću. Otkrio je da kada je jednosmerni krug zatvoren, ispod negativne elektrode, tj. ekscitabilnost se povećava ispod katode, a smanjuje pod pozitivnom anodom. Ovo se zove zakon jednosmerne struje. Promjena ekscitabilnosti tkiva (na primjer, živca) pod djelovanjem jednosmjerne struje u području anode ili katode naziva se fiziološki električni ton. Sada je utvrđeno da se pod djelovanjem negativne elektrode - katode smanjuje potencijal stanične membrane. Ovaj fenomen se naziva fizički katelektroton. Pod pozitivnom - anodom, povećava se. Postoji fizički anelektrton. Pošto se pod katodom membranski potencijal približava kritičnom nivou depolarizacije, povećava se ekscitabilnost ćelija i tkiva. Pod anodom se membranski potencijal povećava i udaljava od kritičnog nivoa depolarizacije, pa se ekscitabilnost ćelije i tkiva smanjuje. Treba napomenuti da uz vrlo kratkotrajno djelovanje istosmjerne struje (1 ms ili manje), MP nema vremena za promjenu, pa se ne mijenja ni ekscitabilnost tkiva ispod elektroda.
Jednosmjerna struja se široko koristi u klinici za liječenje i dijagnostiku. Na primjer, koristi se za električnu stimulaciju živaca i mišića, fizioterapiju: jontoforezu i galvanizaciju.
^ Struktura i funkcije citoplazmatske membrane stanica.
Citoplazmatska ćelijska membrana sastoji se od tri sloja: vanjskog proteinskog sloja, srednjeg bimolekularnog lipidnog sloja i unutrašnjeg proteinskog sloja. Debljina membrane je 7,5-10 nM. Bimolekularni sloj lipida je matriks membrane. Molekuli lipida oba sloja su u interakciji sa molekulima proteina koji su uronjeni u njih. Od 60 do 75% membranskih lipida su fosfolipidi, 15-30% holesterol. Proteini su uglavnom predstavljeni glikoproteinima. Postoje integralni proteini koji prodiru kroz cijelu membranu i periferni proteini koji se nalaze na vanjskoj ili unutrašnjoj površini. Integralni proteini formiraju jonske kanale koji osiguravaju razmjenu određenih jona između ekstra- i intracelularne tekućine. Oni su takođe enzimi koji provode antigradijentni transport jona kroz membranu. Periferni proteini su hemoreceptori na vanjskoj površini membrane, koji mogu stupiti u interakciju s različitim PAS.
Funkcije membrane:
Osigurava integritet ćelije kao strukturne jedinice tkiva.
Obavlja razmjenu jona između citoplazme i ekstracelularne tekućine.
4. Proizvodi percepciju i obradu informacija koje dolaze u ćeliju u obliku hemijskih i električnih signala.
^ Mehanizmi podražljivosti ćelija. Membranski jonski kanali.
Mehanizmi nastanka membranskog potencijala (MP) i akcionog potencijala (AP)
U osnovi, informacije koje se prenose u tijelu su u obliku električnih signala (na primjer, nervnih impulsa). Po prvi put je prisustvo životinjskog elektriciteta utvrdio fiziolog L. Galvani 1786. godine. Da bi proučavao atmosferski elektricitet, okačio je neuromišićne preparate žabljih krakova na bakrenu kuku. Kada su ove šape dotakle željeznu ogradu balkona, mišići su se skupili. To je ukazivalo na djelovanje neke vrste elektriciteta na nerv neuromišićnog preparata. Galvani je smatrao da je to zbog prisustva elektriciteta u samim živim tkivima. Međutim, A. Volta je otkrio da je izvor električne energije mjesto kontakta dva različita metala – bakra i željeza. U fiziologiji se Galvanijevim prvim klasičnim iskustvom smatra dodirivanje živca neuromišićnim preparatom bimetalnom pincetom od bakra i željeza. Da bi dokazao svoj slučaj, Galvani je napravio drugi eksperiment. Bacio je kraj živca koji je inervirao neuromišićni preparat preko posekotine njegovog mišića. Rezultat je bila kontrakcija. Međutim, ovo iskustvo nije uvjerilo Galvanijeve savremenike. Stoga je drugi talijanski Matteuchi napravio sljedeći eksperiment. Nerv jednog neuromišićnog žabljeg preparata je nadgradio na mišić drugog, koji se kontrahirao pod uticajem nadražujuće struje. Kao rezultat toga, prvi lijek je također počeo opadati. To je ukazivalo na prijenos električne energije (PD) s jednog mišića na drugi. Prisustvo razlike potencijala između oštećenih i neoštećenih dijelova mišića prvi put je točno utvrđeno u 19. stoljeću pomoću strunog galvanometra (ampermetra) Matteuchi. Štaviše, rez je imao negativan naboj, a površina mišića pozitivna.
^ Klasifikacija i struktura jonskih kanala u citoplazmi
membrane. Mehanizmi nastanka membranskog potencijala
i akcioni potencijali.
Prvi korak u proučavanju uzroka ekscitabilnosti ćelija napravio je engleski fiziolog Donann u svom djelu The Theory of Membrane Equilibrium 1924. godine. On je teorijski utvrdio da je razlika potencijala unutar ćelije i izvan nje, tj. potencijal mirovanja ili MP, blizu je potencijalu ravnoteže kalijuma. To je potencijal formiran na polupropusnoj membrani koja razdvaja otopine s različitim koncentracijama kalijevih jona, od kojih jedan sadrži velike nepenetrirajuće anione. Nernst je precizirao svoje proračune. Izveo je jednačinu potencijala difuzije. Za kalijum će biti jednako:
Ek=58 lg -------- = 58 lg ----- = - 75 mV,
Ovo je teoretski izračunata vrijednost MP.
Eksperimentalno, mehanizme za nastanak razlike potencijala između ekstracelularne tečnosti i citoplazme, kao i ekscitaciju ćelija, ustanovili su 1939. godine u Kembridžu Hodžkin i Haksli. Ispitivali su džinovsko nervno vlakno (akson) lignje i otkrili da unutarćelijska tečnost neurona sadrži 400 mM kalijuma, 50 mM natrijuma, 100 mM hlora i vrlo malo kalcijuma. Ekstracelularna tečnost je sadržavala samo 10 mM kalijuma, 440 mM natrijuma, 560 mM hlora i 10 mM kalcijuma. Dakle, postoji višak kalijuma unutar ćelija, a natrijuma i kalcijuma izvan njih. To je zbog činjenice da su u ćelijsku membranu ugrađeni jonski kanali koji reguliraju propusnost membrane za jone natrijuma, kalija, kalcija i hlora.
Svi jonski kanali su podijeljeni u sljedeće grupe:
1. Po selektivnosti:
A) Selektivna, tj. specifično. Ovi kanali su propusni za strogo definisane jone.
B) Niska selektivnost, nespecifična, bez određene ionske selektivnosti. U membrani ih je samo nekoliko.
2. Po prirodi prenesenih jona:
A) kalijum
B) natrijum
B) kalcijum
D) hlor
3. Prema stopi inaktivacije, tj. zatvaranje:
A) brzo inaktivirajuće, tj. brzo prelazi u zatvoreno stanje. Oni pružaju brzo rastuće smanjenje MP i isti brzi oporavak.
B) polako se deaktivira. Njihovo otvaranje uzrokuje sporo smanjenje MP i njegov spor oporavak.
Mehanizmi za otvaranje:
B) hemozavisna, otvara se kada su hemoreceptori ćelijske membrane izloženi fiziološki aktivnim supstancama (neurotransmiteri, hormoni itd.).
Sada je utvrđeno da jonski kanali imaju sljedeću strukturu:
1. Selektivni filter koji se nalazi na ušću kanala. Osigurava prolaz strogo određenih jona kroz kanal.
2. Aktivacijska kapija koja se otvara na određenom nivou membranskog potencijala ili djelovanjem odgovarajućeg PAS-a. Aktivacijska kapija naponskih kanala ima senzor koji ih otvara na određenom MP nivou.
3. Inaktivaciona kapija, koja osigurava zatvaranje kanala i prekid provođenja jona kroz kanal na određenom nivou MP (Sl.).
Nespecifični jonski kanali nemaju kapiju.
Selektivni jonski kanali mogu biti u tri stanja, koja su određena pozicijom aktivacijskih (m) i inaktivacionih (h) kapija (sl.):
1.Zatvoreno kada su aktivacijski zatvoreni, a oni za deaktivaciju otvoreni.
2.Aktivirano, obje kapije su otvorene.
3. Deaktivirano, aktivacijske kapije su otvorene, a inaktivacione kapije zatvorene.
Ukupna provodljivost za određeni ion određena je brojem istovremeno otvorenih odgovarajućih kanala. U mirovanju su otvoreni samo kalijevi kanali koji održavaju određeni membranski potencijal, a natrijumski kanali su zatvoreni. Zbog toga je membrana selektivno propusna za kalij, a vrlo malo za jone natrijuma i kalcija, zbog prisustva nespecifičnih kanala. Odnos propusnosti membrane za kalij i natrijum u mirovanju je 1:0,04. Joni kalija ulaze u citoplazmu i akumuliraju se u njoj. Kada njihov broj dostigne određenu granicu, oni počinju napuštati ćeliju kroz otvorene kalijeve kanale duž gradijenta koncentracije. Međutim, oni ne mogu pobjeći s vanjske površine stanične membrane. Tamo ih drži električno polje negativno nabijenih aniona smještenih na unutrašnjoj površini. To su sulfatni, fosfatni i nitratni anioni, anionske grupe aminokiselina za koje je membrana nepropusna. Stoga se pozitivno nabijeni kationi kalija akumuliraju na vanjskoj površini membrane, a negativno nabijeni anioni akumuliraju se na unutrašnjoj površini. Postoji transmembranska razlika potencijala. Rice.
Oslobađanje kalijevih jona iz ćelije se dešava sve dok potencijal koji se pojavljuje sa pozitivnim predznakom izvana ne uravnoteži gradijent koncentracije kalijuma usmerenog van ćelije. One. joni kalija akumulirani na vanjskoj strani membrane neće odbijati iste ione prema unutra. Pojavljuje se određeni membranski potencijal, čiji je nivo određen provodljivošću membrane za jone kalija i natrija u mirovanju. U prosjeku, vrijednost potencijala mirovanja je blizu Nernstovog potencijala ravnoteže kalijuma. Na primjer, MP nervnih ćelija je 55-70 mV, prugastih - 90-100 mV, glatkih mišića - 40-60 mV, žljezdanih ćelija - 20-45 mV. Niža stvarna vrijednost MP ćelija objašnjava se činjenicom da njenu vrijednost smanjuju joni natrijuma, za koje je membrana slabo propusna i mogu ući u citoplazmu. S druge strane, negativni ioni klorida koji ulaze u ćeliju neznatno povećavaju MP.
Budući da je membrana u mirovanju blago propusna za jone natrija, potreban je mehanizam za uklanjanje ovih jona iz ćelije. To je zbog činjenice da bi postepeno nakupljanje natrijuma u ćeliji dovelo do neutralizacije membranskog potencijala i nestanka ekscitabilnosti. Ovaj mehanizam se naziva natrijum-kalijum pumpa. Održava razliku između koncentracija kalija i natrijuma na obje strane membrane. Natrijum-kalijum pumpa je enzim natrijum-kalijum ATPaza. Njegovi proteinski molekuli ugrađeni su u membranu. On razgrađuje ATP i koristi oslobođenu energiju za uklanjanje natrijuma iz ćelije i pumpanje kalija u nju. U jednom ciklusu, svaki molekul natrijum-kalijum ATPaze uklanja 3 jona natrijuma i doprinosi 2 jona kalijuma. Budući da manje pozitivno nabijenih jona ulazi u ćeliju nego što se uklanja iz nje, natrijum-kalijum ATPaza povećava membranski potencijal za 5-10 mV.
Membrana ima sljedeće mehanizme transmembranskog transporta jona i drugih supstanci:
1. Aktivni transport. Izvodi se pomoću energije ATP-a. U ovu grupu transportnih sistema spadaju natrijum-kalijum pumpa, pumpa kalcijuma, pumpa za hlor.
2. Pasivni transport. Kretanje jona se vrši duž gradijenta koncentracije bez potrošnje energije. Na primjer, ulazak kalijuma u ćeliju i izlazak iz nje kroz kalijumove kanale.
3. Povezani transport. Anti-gradijentni transport jona bez potrošnje energije. Na primjer, na ovaj način se odvija jonska izmjena natrijum-natrijum, natrijum-kalcijum, kalijum-kalijum. Nastaje zbog razlike u koncentraciji drugih jona.
Membranski potencijal se snima metodom mikroelektroda. Da bi se to postiglo, tanka, manje od 1 μM staklena mikroelektroda se uvodi kroz membranu u citoplazmu ćelije. Napunjen je fiziološkim rastvorom. Druga elektroda se postavlja u tečnost koja okružuje ćelije. Od elektroda signal ide do biopotencijalnog pojačala, a od njega do osciloskopa i rekordera (sl.).
Dalja istraživanja Hodgkina i Huxleya su pokazala da kada je akson lignje pobuđen, dolazi do brze fluktuacije membranskog potencijala, koja je imala oblik vrha (šiljka) na ekranu osciloskopa. Ovu oscilaciju su nazvali akcionim potencijalom (AP). Budući da je električna struja za ekscitabilne membrane adekvatan stimulans, AP se može inducirati postavljanjem negativne elektrode, katode, na vanjsku površinu membrane, a anode na unutarnjoj pozitivnoj. To će dovesti do smanjenja vrijednosti naboja membrane - njegove depolarizacije. Pod dejstvom slabe struje ispod praga dolazi do pasivne depolarizacije, tj. javlja se katelektroton (sl.). Ako se jačina struje poveća do određene granice, tada će se na kraju perioda njenog utjecaja na katelektronski plato pojaviti mali spontani porast - lokalni ili lokalni odgovor. To je posljedica otvaranja malog dijela natrijumskih kanala koji se nalaze ispod katode. Sa graničnom strujom, MP se smanjuje do kritičnog nivoa depolarizacije (CDL), na kojem počinje stvaranje akcionog potencijala. Za neurone je otprilike na nivou -50 mV.
Na krivulji akcionog potencijala razlikuju se sljedeće faze:
1. Lokalni odgovor (lokalna depolarizacija) koji prethodi razvoju PD.
2. Faza depolarizacije. Tokom ove faze, MF se brzo smanjuje i dostiže nulu. Nivo depolarizacije raste iznad 0. Zbog toga membrana dobija suprotan naboj – unutar nje postaje pozitivna, a izvana negativna. Fenomen promjene naboja membrane naziva se reverzija membranskog potencijala. Trajanje ove faze u nervnim i mišićnim ćelijama je 1-2 ms.
3. Faza repolarizacije. Počinje kada se dostigne određeni nivo MP (približno +20 mV). Potencijal membrane počinje brzo da se vraća na potencijal mirovanja. Trajanje faze 3-5 ms.
4. Faza depolarizacije tragova ili negativni potencijal traga. Period kada se vraćanje MP u potencijal mirovanja privremeno odgađa. Traje 15-30 ms.
5. Faza hiperpolarizacije traga ili pozitivnog potencijala traga. U ovoj fazi, MP neko vrijeme postaje viši od početnog nivoa PP. Njegovo trajanje je 250-300 ms.
Amplituda akcionog potencijala skeletnih mišića je u prosjeku 120-130 mV, neurona 80-90 mV, glatkih mišićnih ćelija 40-50 mV. Kada su neuroni pobuđeni, AP se javlja u početnom segmentu aksona – brežuljku aksona.
Pojava AP je posljedica promjene ionske permeabilnosti membrane nakon ekscitacije. Tokom perioda lokalnog odgovora, spori natrijumski kanali se otvaraju, dok brzi ostaju zatvoreni i dolazi do privremene spontane depolarizacije. Kada MP dostigne kritični nivo, zatvorena aktivaciona kapija natrijumskih kanala se otvaraju i joni natrijuma jure u ćeliju poput lavine, uzrokujući progresivnu depolarizaciju. U ovoj fazi otvaraju se i brzi i spori natrijumski kanali. One. natrijum propusnost membrane se naglo povećava. Štaviše, vrijednost kritičnog nivoa depolarizacije ovisi o osjetljivosti aktivacijskih, što je veća, to je niži FCA i obrnuto.
Kada se količina depolarizacije približi potencijalu ravnoteže za jone natrija (+20 mV). jačina gradijenta koncentracije natrijuma je značajno smanjena. Istovremeno počinje proces inaktivacije brzih natrijevih kanala i smanjenje natrijeve provodljivosti membrane. Depolarizacija prestaje. Izlaz kalijevih jona se naglo povećava, tj. izlazna struja kalijuma. U nekim ćelijama to je zbog aktivacije posebnih kanala za izlaz kalija. Ova struja, usmjerena iz ćelije, služi za brzo prebacivanje MP na nivo potencijala mirovanja. One. počinje faza repolarizacije. Povećanje MP dovodi do zatvaranja aktivacijskih kapija natrijumskih kanala, što dodatno smanjuje natrijumsku permeabilnost membrane i ubrzava repolarizaciju.
Pojava faze depolarizacije u tragovima se objašnjava činjenicom da ne večina spori natrijumski kanali ostaju otvoreni.
Hiperpolarizacija u tragovima je povezana sa povećanom, nakon PD, kalijumom provodljivosti membrane i činjenicom da je natrijum-kalijum pumpa aktivnija, izvodeći ione natrijuma koji su ušli u ćeliju tokom PD.
Promjenom provodljivosti brzih natrijumovih i kalijumovih kanala moguće je uticati na stvaranje AP, a time i na pobuđivanje ćelija. Uz potpunu blokadu natrijevih kanala, na primjer, s otrovom tetrodontne ribe - tetrodotoksinom, stanica postaje neuzbudljiva. Ovo se koristi u klinici. Takve lokalni anestetici, jer novokain, dikain, lidokain inhibiraju tranziciju natrijumskih kanala nervnih vlakana u otvoreno stanje. Zbog toga prestaje provođenje nervnih impulsa duž senzornih nerava, dolazi do anestezije (anestezije) organa. Uz blokadu kalijevih kanala, otežano je oslobađanje kalijevih jona iz citoplazme na vanjsku površinu membrane; MP oporavak. Zbog toga se faza repolarizacije produžava. Ovaj efekat blokatora kalijumovih kanala se takođe koristi u kliničkoj praksi. Na primjer, jedan od njih, kinidin, produžava fazu repolarizacije kardiomiocita, usporava srčane kontrakcije i normalizira srčani ritam.
Takođe treba napomenuti da što je veća brzina širenja AP duž membrane ćelije ili tkiva, to je veća i njena provodljivost.
^ Odnos faza akcionog potencijala i ekscitabilnosti
Nivo ćelijske ekscitabilnosti zavisi od AP faze. U fazi lokalnog odgovora, ekscitabilnost se povećava. Ova faza ekscitabilnosti naziva se latentno dodavanje.
U fazi repolarizacije AP, kada se otvore svi natrijumski kanali i natrijum joni jure u ćeliju poput lavine, čak ni superjaki stimulans ne može da stimuliše ovaj proces. Dakle, faza depolarizacije odgovara fazi potpune neekscitabilnosti ili apsolutne refraktornosti.
Tokom faze repolarizacije zatvara se sve više i više natrijumovih kanala. Međutim, mogu se ponovo otvoriti pod dejstvom stimulusa iznad praga. One. razdražljivost ponovo počinje rasti. Ovo odgovara fazi relativne neekscitabilnosti ili relativne refraktornosti.
Tokom depolarizacije u tragovima, MP je na kritičnom nivou, tako da čak i stimulansi pre praga mogu izazvati ekscitaciju ćelije. Stoga je u ovom trenutku njena razdražljivost povećana. Ova faza se naziva faza egzaltacije ili natprirodne ekscitabilnosti.
U trenutku hiperpolarizacije traga, MP je veći od početnog nivoa, tj. dalje KUD i njegova ekscitabilnost je smanjena. Ona je u fazi subnormalne ekscitabilnosti. Rice. Treba napomenuti da je fenomen akomodacije povezan i sa promjenom provodljivosti jonskih kanala. Ako se depolarizirajuća struja sporo povećava, onda to dovodi do djelomične inaktivacije natrijuma i aktivacije kalijevih kanala. Stoga ne dolazi do razvoja PD.
^ FIZIOLOGIJA MIŠIĆA
U tijelu postoje 3 vrste mišića: skeletni ili prugasti, glatki i srčani. Skeletni mišići omogućavaju kretanje tijela u prostoru, održavajući držanje tijela zahvaljujući tonusu mišića udova i tijela. Glatki mišići su neophodni za peristaltiku gastrointestinalnog trakta, urinarnog sistema regulacija tonusa krvnih sudova, bronhija itd. Srčani mišić se koristi za kontrakciju srca i pumpanje krvi. Svi mišići imaju ekscitabilnost, provodljivost i kontraktilnost, a srčani i mnogi glatki mišići imaju automatizam - sposobnost spontanih kontrakcija.
^ Ultrastruktura skeletnih mišićnih vlakana.
Motorne jedinice Glavni morfo-funkcionalni element neuromišićnog aparata skeletnih mišića je motorna jedinica. Uključuje motorni neuron kičmene moždine s mišićnim vlaknima inerviranim njegovim aksonom. Unutar mišića, ovaj akson formira nekoliko terminalnih grana. Svaka takva grana formira kontakt - neuromišićnu sinapsu na zasebnom mišićnom vlaknu. Nervni impulsi koji dolaze iz motornog neurona uzrokuju kontrakcije određene grupe mišićnih vlakana.
Skeletni mišići se sastoje od mišićnih snopova formiranih od velikog broja mišićnih vlakana. Svako vlakno je cilindrična ćelija prečnika 10-100 mikrona i dužine od 5 do 400 mikrona. Ima ćelijsku membranu - sarkolemu. Sarcoplazma sadrži nekoliko jezgara, mitohondrija, formacije sarkoplazmatskog retikuluma (SR) i kontraktilne elemente - miofibrile. Sarkoplazemski retikulum ima osebujnu strukturu. Sastoji se od sistema poprečnih, uzdužnih cijevi i rezervoara. Poprečni tubuli su produžeci sarkoplazme u ćeliju. Na njih se spajaju uzdužne cijevi sa vodokotlićima. Zbog toga se akcioni potencijal može širiti iz sarkoleme u sistem sarkoplazmatskog retikuluma. Mišićno vlakno sadrži više od 1000 miofibrila smještenih duž njega. Svaka miofibrila se sastoji od 2500 protofibrila ili miofilamenata. To su filamenti kontraktilnih proteina aktina i miozina. Protofibrile miozina su debele, aktinske protofibrile su tanke.
Na miozinskim filamentima nalaze se poprečni nastavci sa glavama koje se pružaju pod uglom. U vlaknima skeletnih mišića svjetlosna mikroskopija pokazuje poprečnu prugastost, tj. naizmjenično svijetle i tamne pruge. Tamne trake se nazivaju A-diskovi ili anizotropni, svijetli I-diskovi (izotropni). Miozinski filamenti su koncentrisani u A-diskovima, koji imaju anizotropiju i stoga imaju tamnu boju. I-diskovi su formirani aktinskim filamentima. Tanka Z-ploča je vidljiva u sredini I-diskova. Za njega su vezani protofibrili aktina. Dio miofibrila između dvije Z-lamele naziva se sarkomer. To je strukturni element miofibrila. U mirovanju, debeli miozinski filamenti ulaze u praznine između aktinskih filamenata samo na kratkoj udaljenosti. Stoga se u srednjem dijelu A-diska nalazi svjetlija H-zona, gdje nema aktinskih filamenata. Elektronska mikroskopija pokazuje veoma tanku M-liniju u njenom centru. Formiran je od lanaca potpornih proteina za koje su vezani protofibrili miozina (Sl.
^ Mehanizmi mišićne kontrakcije
Svjetlosnom mikroskopijom uočeno je da se u trenutku kontrakcije širina A-diska ne smanjuje, ali se I-diskovi i H-zone sarkomera sužavaju. Elektronskim mikroskopom je utvrđeno da se dužina aktinskih i miozinskih filamenata ne mijenja u trenutku kontrakcije. Tako su Huxley i Hanson razvili teoriju klizanja niti. Prema njemu, mišić se skraćuje kao rezultat kretanja tankih aktinskih filamenata u prazninama između miozinskih filamenata. To dovodi do skraćivanja svakog sarkomera koji formira miofibrile. Klizanje filamenata nastaje zbog činjenice da su, pri prelasku u aktivno stanje, glave miozinskih procesa povezane sa centrima aktinskih filamenata i uzrokuju njihovo pomicanje u odnosu na sebe (pokreti). Ali ovo je posljednja faza cijelog kontraktilnog mehanizma. Kontrakcija počinje činjenicom da se PD javlja u području završne ploče motornog živca. Širi se velikom brzinom duž sarkoleme i od nje prelazi duž sistema poprečnih tubula SR do uzdužnih tubula i cisterni. Dolazi do depolarizacije membrane rezervoara i ioni kalcija se oslobađaju iz njih u sarkoplazmu. Aktinski filamenti sadrže molekule još dva proteina - troponina i tropomiozina. Pri niskoj (manje od 10 -8 M) koncentraciji kalcija, tj. u mirovanju, tropomiozin blokira vezivanje miozinskih mostova za aktinske filamente. Kada ioni kalcija počnu napuštati SR, molekula troponina mijenja svoj oblik na takav način da oslobađa aktinske aktivne centre od tropomiozina. Glave miozina su pričvršćene za ove centre i klizanje počinje zbog ritmičkog vezivanja i razdvajanja poprečnih mostova aktinskim filamentima. U ovom slučaju, glave se ritmično kreću duž aktinskih filamenata do Z-membrana. Potrebno je 50 takvih ciklusa da bi se mišić u potpunosti kontrahirao. Prijenos signala od pobuđene membrane do miofibrila naziva se elektromehaničko spajanje. Kada prestane stvaranje AP i membranski potencijal se vrati na prvobitni nivo, Ca-pumpa (enzim Ca-ATPaza) počinje da radi. Kalcijumovi joni se ponovo upumpavaju u cisterne sarkoplazmatskog retikuluma i njihova koncentracija pada ispod 10 -8 M. Molekuli troponina dobijaju svoj prvobitni oblik i tropomiozin ponovo počinje da blokira aktivne centre aktina. Glave miozina se odvajaju od njih i mišić se vraća u prvobitno opušteno stanje zbog elastičnosti.
^ Energija mišićne kontrakcije
ATP je izvor energije za kontrakciju i opuštanje. Glave miozina imaju katalitička mjesta koja razgrađuju ATP do ADP i neorganskog fosfata. One. miozin je takođe enzim ATPaze. Aktivnost miozina kao ATPaze značajno se povećava kada je u interakciji s aktinom. Sa svakim ciklusom interakcije između aktina i glave, miozin dijeli 1 ATP molekul. Stoga, što više mostova prelazi u aktivno stanje, što se ATP više dijeli, to je jača kontrakcija. Da bi se stimulirala ATPazna aktivnost miozina, potrebni su joni kalcija, koji se oslobađaju iz SR, koji potiču oslobađanje aktinskih aktivnih centara iz tropomiozina. Međutim, opskrba ATP-om u ćeliji je ograničena. Stoga se za obnavljanje rezervi ATP-a obnavlja - resinteza. Izvodi se anaerobno i aerobno. Proces anaerobne resinteze provode fosfageni i glikolitički sistemi. Prvi koristi rezerve kreatin fosfata za obnavljanje ATP-a. Razgrađuje se na kreatin i fosfat, koji se uz pomoć enzima (ADP + P = ATP) prenosi na ADP.Sistem fosfagene resinteze daje najveću moć kontrakcije, ali zbog male količine kreatin fosfata u ćeliji, funkcionira samo za 5-6 sekundi kontrakcije. Glikolitički sistem koristi anaerobnu razgradnju glukoze (glikogena) do mliječne kiseline za resintezu ATP-a. Svaki molekul glukoze osigurava oporavak tri ATP molekula. Energetski potencijal ovog sistema je veći od fosfagenog, ali i on može poslužiti kao izvor energije kontrakcije samo 0,5 - 2 minuta. Istovremeno, rad glikolitičkog sistema prati nakupljanje mliječne kiseline u mišićima i smanjenje sadržaja kisika. Uz produženi rad, uz pojačanu cirkulaciju krvi, počinje se odvijati resinteza ATP-a uz pomoć oksidativne fosforilacije, tj. aerobno. Energetski potencijal oksidacionog sistema je mnogo veći od ostalih. Proces nastaje zbog oksidacije ugljikohidrata i masti. Intenzivnim radom uglavnom se oksidiraju ugljikohidrati, a umjerenim radom masti. Za opuštanje je potrebna i ATP energija. Nakon smrti, sadržaj ATP-a u ćelijama se brzo smanjuje, a kada padne ispod kritičnog nivoa, miozinski poprečni mostovi se ne mogu odvojiti od aktinskih filamenata (sve do enzimske autolize ovih proteina). Pojavljuje se mortis ukočenost. ATP je neophodan za opuštanje jer održava Ca-pumpu u radu.
^ Biomehanika mišićnih kontrakcija.
Pojedinačna kontrakcija, sumacija, tetanus.
Kada se iritacija jednog praga ili iznad praga primeni na motorni nerv ili mišić, dolazi do jedne kontrakcije. Svojom grafičkom registracijom na rezultujućoj krivulji mogu se razlikovati tri uzastopna perioda:
1. Latentni period. Ovo je vrijeme od trenutka kada je iritacija primijenjena do početka kontrakcije. Njegovo trajanje je oko 1-2 ms. Tokom latentnog perioda, AP se stvara i razmnožava, kalcijum se oslobađa iz SR, aktin stupa u interakciju sa miozinom, itd.
2. Period skraćivanja. U zavisnosti od tipa mišića (brzi ili spori), njegovo trajanje je od 10 do 100 ms.,
3. Period opuštanja. Njegovo trajanje je nešto duže od skraćivanja. Rice.
U načinu jedne kontrakcije, mišić je sposoban da radi dugo bez umora, ali je njegova snaga beznačajna. Stoga su takve kontrakcije rijetke u tijelu, na primjer, brzi okulomotorički mišići mogu se kontrahirati na ovaj način. Češće se zbrajaju pojedinačne kontrakcije.
Sumiranje je dodavanje 2 uzastopne kontrakcije kada se na njega primjenjuju 2 praga ili suprapraga podražaja, interval između kojih je kraći od trajanja jedne kontrakcije, ali duži od trajanja refraktornog perioda. Postoje dvije vrste zbrajanja: potpuno i nepotpuno. Nepotpuno zbrajanje nastaje ako se ponovljena stimulacija primjenjuje na mišić kada se već počeo opuštati. Potpuna nastaje kada ponovljena iritacija djeluje na mišić prije početka perioda opuštanja, tj. na kraju perioda skraćivanja (slika 1.2). Amplituda kontrakcije kod punog zbrajanja je veća nego kod nepotpunog. Ako se interval između dvije iritacije dodatno smanji. Na primjer, primijenite drugi usred perioda skraćivanja, tada neće biti sumiranja, jer je mišić u refraktornom stanju.
Tetanus- ovo je duga kontrakcija mišića koja je rezultat zbrajanja nekoliko pojedinačnih kontrakcija koje se razvijaju kada se na njega primijeni niz uzastopnih podražaja. Postoje 2 oblika tetanusa: nazubljeni i glatki. Nazubljeni tetanus se opaža ako svaka sljedeća iritacija djeluje na mišić kada je već počeo da se opušta. One. uočeno je nepotpuno zbrajanje (sl.). Glatki tetanus nastaje kada se svaki sljedeći stimulans primjenjuje na kraju perioda skraćivanja. One. postoji potpuni zbir pojedinačnih kontrakcija i (sl.). Amplituda glatkog tetanusa je veća od amplituda nazubljenog. Normalno, ljudski mišići se kontrahuju u glatkom modusu tetanusa. Nazubljen se javlja kod patologije, kao što je tremor ruku uz intoksikaciju alkoholom i Parkinsonova bolest.
^ Utjecaj učestalosti i jačine stimulacije na amplitudu kontrakcije
Ako postupno povećavate učestalost iritacije, tada se povećava amplituda tetaničke kontrakcije. Na određenoj frekvenciji, ona će postati maksimalna. Ova frekvencija se naziva optimalnom. Daljnje povećanje učestalosti iritacije praćeno je smanjenjem snage.
Tetanička kontrakcija. Frekvencija na kojoj amplituda kontrakcije počinje da se smanjuje naziva se pesimalna frekvencija. Pri vrlo visokoj frekvenciji stimulacije, mišić se ne kontrahira (sl.). Koncept optimalnih i pesimalnih frekvencija predložio je N.E. Vvedensky. Otkrio je da svaka stimulacija granične ili supragranične sile, koja uzrokuje kontrakciju, istovremeno mijenja ekscitabilnost mišića. Stoga se postupnim povećanjem frekvencije stimulacije djelovanje impulsa sve više pomjera na početak perioda opuštanja, tj. faza egzaltacije. Na optimalnoj frekvenciji svi impulsi djeluju na mišić u fazi egzaltacije, tj. povećana razdražljivost. Stoga je amplituda tetanusa maksimalna. Daljnjim povećanjem frekvencije stimulacije, sve veći broj impulsa djeluje na mišić u refraktornoj fazi. Amplituda tetanusa se smanjuje.
Jedno mišićno vlakno, kao i svaka podražljiva ćelija, reaguje na iritaciju prema zakonu sve ili ništa. Mišić se povinuje zakonu sile. Sa povećanjem snage stimulacije, povećava se i amplituda njegove kontrakcije. Pri određenoj (optimalnoj) sili, amplituda postaje maksimalna. Međutim, ako se jačina stimulacije dodatno poveća, amplituda kontrakcije se ne povećava, već čak i smanjuje zbog katodne depresije. Takva sila bi bila pesimalna. Takva reakcija mišića objašnjava se činjenicom da se sastoji od vlakana različite ekscitabilnosti, pa je povećanje snage iritacije praćeno ekscitacijom sve većeg broja njih. Pri optimalnoj snazi sva vlakna su uključena u kontrakciju. Katodna depresija je smanjenje ekscitabilnosti pod djelovanjem depolarizirajuće struje - katode, velike sile ili trajanja.
^ načini smanjenja. Snaga i rad mišića.
Postoje sljedeći načini mišićne kontrakcije:
1. Izotonične kontrakcije. Dužina mišića se smanjuje, ali se tonus ne mijenja. Nisu uključeni u motoričke funkcije tijela.
2.Izometrijska kontrakcija. Dužina mišića se ne mijenja, ali se tonus povećava. Osnovni statički rad, kao što je održavanje držanja tijela.
3. Auksotonične kontrakcije. Dužina i tonus mišića se također mijenjaju. Uz pomoć njih dolazi do kretanja tijela, drugih motoričkih radnji.
Maksimalna mišićna snaga je količina maksimalne napetosti koju mišić može razviti. Zavisi od strukture mišića, njegovog funkcionalnog stanja, početne dužine, pola, starosti, stepena treniranosti osobe.
Ovisno o građi, razlikuju se mišići s paralelnim vlaknima (na primjer, krojački), fusiformni (biceps brachii), perasti (teleni). Ove vrste mišića imaju različitu fiziološku površinu poprečnog presjeka. To je zbir površina poprečnog presjeka svih mišićnih vlakana koja čine mišić. Najveća fiziološka površina poprečnog presjeka, a samim tim i snaga, nalazi se u perastim mišićima. Najmanji u mišićima sa paralelnim rasporedom vlakana (sl.).
S umjerenim istezanjem mišića povećava se sila njegove kontrakcije, ali se s prekomjernim istezanjem smanjuje. Uz umjereno zagrijavanje, također se povećava, a opada sa hlađenjem. Snaga mišića opada zbog umora, metaboličkih poremećaja itd. .Maksimalna snaga raznih mišićnih grupa se određuje dinamometrima, ručnim dizanjem, mrtvim dizanjem itd.
Da bi se uporedile snage različitih mišića, određuje se njihova specifična ili apsolutna snaga. Ona je jednaka maksimumu podijeljenom sa sq. vidi površinu poprečnog presjeka mišića. Specifična snaga ljudskog gastrocnemius mišića je 6,2 kg/cm2, mišića tricepsa je 16,8 kg/cm2, a žvačnog mišića 10 kg/cm2.
Rad mišića se dijeli na dinamički i statički.Dinamički se izvodi kada se opterećenje pomjera. Tokom dinamičkog rada mijenja se dužina mišića i njegova napetost. Stoga mišić radi u auksotoničnom režimu. At statički rad nema kretanja tereta, tj. mišić radi u izometrijskom režimu. Dinamički rad jednak je proizvodu težine tereta i visine njegovog uspona ili količini skraćivanja mišića (A = P * h). Rad se mjeri u kg.m, džulima. Ovisnost količine rada od opterećenja podliježe zakonu prosječnih opterećenja. Kada se opterećenje poveća, u početku se povećava rad mišića. Pri srednjim opterećenjima postaje maksimalno. Ako se povećanje opterećenja nastavi, onda se rad smanjuje (sl.). Isti uticaj na veličinu dela ima i svoj ritam. Maksimalni rad mišića izvodi se u prosječnom ritmu. Od posebne važnosti u izračunavanju veličine opterećenja je definicija mišićne snage. Ovo je rad obavljen u jedinici vremena (P = A * T). uto
^ Umor mišića
Umor je privremeno smanjenje mišićnih performansi kao rezultat rada. Umor izolovanog mišića može biti uzrokovan njegovom ritmičkom stimulacijom. Kao rezultat, sila kontrakcija progresivno opada (Sl.). Što je veća učestalost, jačina iritacije, veličina opterećenja, brže se razvija umor. Sa umorom, krivulja jedne kontrakcije značajno se mijenja. Trajanje latentnog perioda, perioda skraćivanja, a posebno perioda opuštanja se povećava, ali se amplituda smanjuje (Sl.). Što je zamor mišića jači, to je duže trajanje ovih perioda. U nekim slučajevima ne dolazi do potpunog opuštanja. Razvija se kontraktura. Ovo je stanje produžene nevoljne kontrakcije mišića. Rad mišića i umor se ispituju ergografijom.
U prošlom veku, na osnovu eksperimenata sa izolovanim mišićima, predložene su 3 teorije zamora mišića.
1. Schiffova teorija: umor je posljedica iscrpljivanja energetskih rezervi u mišićima.
2. Pflugerova teorija: umor je posljedica nakupljanja metaboličkih produkata u mišićima.
3. Verworn-ova teorija: umor je posljedica nedostatka kisika u mišićima.
Zaista, ovi faktori doprinose umoru u eksperimentima na izolovanim mišićima. U njima je poremećena resinteza ATP-a, nakupljaju se mliječna i pirogrožđana kiselina, sadržaj kisika je nedovoljan. Međutim, u tijelu intenzivno rade mišići primaju neophodan kisik, hranjive tvari i oslobađaju se od metabolita zbog pojačane opće i regionalne cirkulacije krvi. Stoga su predložene druge teorije zamora. Konkretno, neuromuskularne sinapse igraju određenu ulogu u umoru. Umor u sinapsi nastaje zbog iscrpljivanja zaliha neurotransmitera. Međutim, glavna uloga u zamoru motoričkog aparata pripada motornim centrima centralnog nervnog sistema. U prošlom vijeku I.M. Sechenov je ustanovio da ako se mišići jedne ruke zamaraju, onda se njihov učinak brže obnavlja kada se radi s drugom rukom ili nogama. Vjerovao je da je to zbog prebacivanja procesa ekscitacije iz jednog motornog centra u drugi. Odmor uz uključivanje drugih mišićnih grupa nazvao je aktivnim. Sada je utvrđeno da je motorički zamor povezan sa inhibicijom odgovarajućih nervnih centara, kao rezultat metaboličkih procesa u neuronima, pogoršanja sinteze neurotransmitera i inhibicije sinaptičke transmisije.
^ motorne jedinice
Glavni morfo-funkcionalni element neuromišićnog aparata skeletnih mišića je motorna jedinica (MU). Uključuje motorni neuron kičmene moždine s mišićnim vlaknima inerviranim njegovim aksonom. Unutar mišića, ovaj akson formira nekoliko terminalnih grana. Svaka takva grana formira kontakt - neuromišićnu sinapsu na zasebnom mišićnom vlaknu. Nervni impulsi koji dolaze iz motornog neurona uzrokuju kontrakcije određene grupe mišićnih vlakana. Motorne jedinice malih mišića koje izvode fine pokrete (mišići oka, ruke) sadrže malu količinu mišićnih vlakana. U velikim ih ima stotine puta više. Sve DU u zavisnosti od funkcionalne karakteristike dijele se u 3 grupe:
I. Sporo neumoran. Formiraju ih "crvena" mišićna vlakna, u kojima ima manje miofibrila. Brzina kontrakcije i snaga ovih vlakana su relativno male, ali nisu jako zamorne. Stoga se nazivaju tonicima. Regulaciju kontrakcija takvih vlakana provodi mali broj motornih neurona, čiji aksoni imaju nekoliko terminalnih grana. Primjer je mišić soleus.
IIB. Brzo, lako se zamara. Mišićna vlakna sadrže mnogo miofibrila i nazivaju se "bijela". Brzo se skupite i razvijajte veliku snagu, ali se brzo umarajte. Stoga se nazivaju fazama. Motorni neuroni ovih MU su najveći, imaju debeli akson sa brojnim terminalnim granama. Generiraju nervne impulse visoke frekvencije. Mišići oka.
IIA. Brz, otporan na zamor. Zauzimaju srednju poziciju.
^ Fiziologija glatkih mišića
Glatki mišići nalaze se u zidovima većine organa za varenje, krvnim sudovima, izvodnim kanalima različitih žlijezda i mokraćnom sistemu. Oni su nevoljni i obezbjeđuju peristaltiku probavnog i urinarnog sistema, održavajući vaskularni tonus. Za razliku od skeletnih, glatke mišiće formiraju ćelije češće vretenaste i male veličine, koje nemaju poprečne pruge. Potonje je zbog činjenice da kontraktilni aparat nema uređenu strukturu. Miofibrile se sastoje od tankih filamenata aktina koji se kreću u različitim smjerovima i pričvršćuju se za različite dijelove sarkoleme. Protofibrili miozina nalaze se pored aktina. Elementi sarkoplazmatskog retikuluma ne formiraju sistem tubula. Odvojene mišićne ćelije su međusobno povezane kontaktima sa niskim električnim otporom - neksusima, što osigurava širenje ekscitacije kroz strukturu glatkih mišića. Ekscitabilnost i provodljivost glatkih mišića je niža od onih skeletnih.
Potencijal membrane je 40-60 mV, budući da MMC membrana ima relativno visoku permeabilnost za jone natrija. Štaviše, u mnogim glatkim mišićima MP nije konstantan. Povremeno se smanjuje i ponovo se vraća na prvobitni nivo. Takve oscilacije se nazivaju spori valovi (SW). Kada vrh sporog talasa dostigne kritični nivo depolarizacije, na njemu počinju da se stvaraju akcioni potencijali, praćeni kontrakcijama (Sl.). MV i PD se provode kroz glatke mišiće brzinom od samo 5 do 50 cm/sek. Takvi glatki mišići nazivaju se spontano aktivnim, tj. oni su automatski. Na primjer, zbog takve aktivnosti dolazi do crijevne peristaltike. Pejsmejkeri crijevne peristaltike nalaze se u početnim dijelovima odgovarajućih crijeva.
Generisanje AP u SMC je posledica ulaska jona kalcijuma u njih. Mehanizmi elektromehaničkog spajanja su također različiti. Kontrakcija se razvija usled ulaska kalcijuma u ćeliju tokom PD Najvažniji ćelijski protein, kalmodulin, posreduje u odnosu kalcijuma sa skraćivanjem miofibrila.
Kriva kontrakcije je također različita. Latentni period, period skraćivanja, a posebno opuštanja, mnogo je duži od perioda skeletnih mišića. Kontrakcija traje nekoliko sekundi. Glatke mišiće, za razliku od skeletnih mišića, karakterizira fenomen plastičnog tonusa. Ova sposobnost je dugo vremena u stanju smanjenja bez značajnije potrošnje energije i umora. Zahvaljujući ovoj osobini, održava se oblik unutrašnjih organa i vaskularni tonus. Osim toga, same ćelije glatkih mišića su receptori za istezanje. Kada se istežu, AP-ovi počinju da se stvaraju, što dovodi do smanjenja SMC-a. Ovaj fenomen se naziva miogeni mehanizam regulacije kontraktilne aktivnosti.