Šta je ćelijska membrana i njene funkcije. Istorija otkrića i proučavanja ćelijske membrane. Plazma membrana, struktura, funkcije
Izvana je ćelija prekrivena plazma membranom (ili vanjskom ćelijskom membranom) debljine oko 6-10 nm.
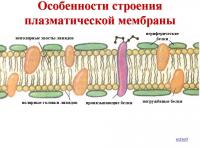
Stanična membrana je gust film proteina i lipida (uglavnom fosfolipida). Molekuli lipida su raspoređeni na uredan način - okomito na površinu, u dva sloja, tako da su njihovi dijelovi koji intenzivno komuniciraju s vodom (hidrofilni) usmjereni prema van, a dijelovi koji su inertni prema vodi (hidrofobni) usmjereni su prema unutra.
Molekuli proteina nalaze se u nekontinuiranom sloju na površini lipidnog okvira s obje strane. Neki od njih su uronjeni u lipidni sloj, a neki prolaze kroz njega, formirajući područja propusna za vodu. Ovi proteini rade razne funkcije- neki od njih su enzimi, drugi su transportni proteini uključeni u prijenos određenih tvari iz okoline u citoplazmu i u obrnuti smjer.
Osnovne funkcije ćelijske membrane
Jedno od glavnih svojstava bioloških membrana je selektivna permeabilnost (semipermeabilnost)- neke tvari prolaze kroz njih otežano, druge lako, pa čak i prema višoj koncentraciji.Tako je za većinu ćelija koncentracija Na iona unutra mnogo niža nego u okruženje. Za K ione karakterističan je obrnuti omjer: njihova koncentracija unutar ćelije je veća nego izvan nje. Zbog toga joni Na uvek teže da uđu u ćeliju, a K joni - da izađu van. Izjednačavanje koncentracija ovih jona sprečeno je prisustvom u membrani posebnog sistema koji ima ulogu pumpe koja pumpa Na ione iz ćelije i istovremeno pumpa K ione unutra.
Želja Na iona da se kreću izvana ka unutra koristi se za transport šećera i aminokiselina u ćeliju. Aktivnim uklanjanjem Na iona iz ćelije stvaraju se uslovi za ulazak glukoze i aminokiselina u nju.
U mnogim ćelijama apsorpcija supstanci se odvija i fagocitozom i pinocitozom. At fagocitoza fleksibilna vanjska membrana formira malu depresiju gdje zarobljena čestica ulazi. Ovo udubljenje se povećava i, okružena dijelom vanjske membrane, čestica je uronjena u citoplazmu ćelije. Fenomen fagocitoze karakterističan je za amebe i neke druge protozoe, kao i za leukocite (fagocite). Slično, postoji apsorpcija od strane ćelija tečnosti koje sadrže potrebna ćeliji supstance. Ovaj fenomen je nazvan pinocitoza.
Spoljne membrane različitih ćelija značajno se razlikuju kako po hemijskom sastavu njihovih proteina i lipida, tako i po njihovom relativnom sadržaju. Upravo te karakteristike određuju raznolikost fiziološke aktivnosti membrana različitih stanica i njihovu ulogu u životu stanica i tkiva.
Endoplazmatski retikulum ćelije povezan je sa spoljašnjom membranom. Uz pomoć vanjskih membrana, Razne vrste međućelijski kontakti, tj. komunikacija između pojedinačnih ćelija.
Mnoge vrste ćelija karakteriše prisustvo na njihovoj površini veliki broj izbočine, nabori, mikroresice. Oni doprinose kako značajnom povećanju površine ćelija i poboljšanju metabolizma, tako i čvršćoj povezanosti pojedinačnih ćelija među sobom.
Na vanjskoj strani ćelijske membrane, biljne stanice imaju debele membrane koje su jasno vidljive u optičkom mikroskopu, a koje se sastoje od celuloze (celuloze). Oni stvaraju jaku potporu biljnim tkivima (drvo).
Neke ćelije životinjskog porijekla imaju i brojne vanjske strukture koje se nalaze na vrhu ćelijske membrane i imaju zaštitni karakter. Primjer je hitin integumentarnih stanica insekata.
Funkcije ćelijske membrane (ukratko)
| Funkcija | Opis |
|---|---|
| zaštitna barijera | Odvaja unutrašnje organele ćelije od spoljašnje okruženje |
| Regulatorno | Reguliše razmenu supstanci između unutrašnjeg sadržaja ćelije i spoljašnje sredine. |
| Razgraničenje (kompartmentalizacija) | Razdvajanje unutrašnjeg prostora ćelije u nezavisne blokove (odjeljke) |
| Energija | - Akumulacija i transformacija energije; - svjetlosne reakcije fotosinteze u hloroplastima; - Apsorpcija i sekrecija. |
| Receptor (informacije) | Učestvuje u formiranju pobude i njenom vođenju. |
| Motor | Obavlja kretanje ćelije ili njenih pojedinačnih delova. |
Glavne ćelijske membrane:
plazma membrana
Plazma membrana koja okružuje svaku ćeliju određuje njenu veličinu, osigurava transport malih i velikih molekula iz ćelije u ćeliju i održava razliku u koncentraciji jona na obje strane membrane. Membrana sudjeluje u međućelijskim kontaktima, percipira, pojačava i prenosi signale iz vanjskog okruženja u ćeliju. Membrana je povezana s mnogim enzimima koji kataliziraju biokemijske reakcije.
nuklearna membrana
Nuklearni omotač se sastoji od vanjske i unutrašnje nuklearne membrane. Nuklearna membrana ima pore kroz koje RNA prodiru iz jezgre u citoplazmu, a regulatorni proteini iz citoplazme u jezgro.
Unutrašnja nuklearna membrana sadrži specifične proteine koji imaju mjesta vezivanja za glavne polipeptide nuklearnog matriksa - lamin A, lamin B i lamin C. Važna funkcija ovih proteina je dezintegracija nuklearne membrane tokom mitoze.
Endoplazmatski retikulum (ER) membrana
ER membrana ima brojne nabore i nabore. Formira kontinuiranu površinu koja ograničava unutrašnji prostor, nazvan ER šupljina. Grubi ER je povezan sa ribozomima, na kojima se sintetišu proteini plazma membrane, ER, Golgijevog aparata, lizozoma i izlučenih proteina. Regije ER koje ne sadrže ribozome nazivaju se glatki ER. Ovdje je završna faza biosinteze kolesterola, fosfolipida, reakcija oksidacije vlastitih metabolita i stranih tvari uz sudjelovanje membranskih enzima - citokrom P 450, citokrom P 450 reduktaza, citokrom b 5 reduktaza i citokrom b 5
golgijev aparat
Golgijev aparat je važna membranska organela odgovorna za modifikaciju, akumulaciju, sortiranje i usmjeravanje različitih supstanci u odgovarajuće unutarćelijske odjeljke, kao i izvan ćelije. Specifični enzimi membrane Golgijevog kompleksa, glikoziltransferaza, glikozilirajući proteini na ostacima serina, treonina ili amidne grupe asparagina, dovršavaju formiranje kompleksnih proteina - glikoproteina.
Mitohondrijalne membrane
Mitohondrije su dvomembranske organele specijalizirane za sintezu ATP-a oksidativnom fosforilacijom. Posebnost vanjske mitohondrijalne membrane je sadržaj velike količine proteina porina, koji formira pore u membrani. Zbog porina, vanjska membrana je slobodno propusna za neorganske ione, metabolite, pa čak i male proteinske molekule (manje od 10 kD). Za velike proteine, vanjska membrana je nepropusna, što omogućava mitohondrijima da zadrže proteine u međumembranskom prostoru od curenja u citosol.
Unutrašnju membranu mitohondrija karakteriše visok sadržaj proteina, oko 70%, koji obavljaju uglavnom katalitičke i transportne funkcije. Membranske translokaze omogućavaju selektivni prijenos tvari iz intermembranskog prostora u matriks i obrnuto; enzimi su uključeni u transport elektrona (lanac transporta elektrona) i sintezu ATP-a.
Membrana lizosoma
Membrana lizosoma ima ulogu "štita" između aktivnih enzima (više od 50), koji pružaju reakcije za razgradnju proteina, ugljikohidrata, masti, nukleinske kiseline, i ostatak sadržaja ćelije. Membrana sadrži jedinstvene proteine, na primjer, ATP zavisnu protonsku pumpu (pumpu), koja održava kiselu sredinu (pH 5) neophodnu za djelovanje hidrolitičkih enzima (proteaza, lipaza), kao i transportne proteine koji omogućavaju razgradnju makromolekula proizvodi koji napuštaju lizozom. Takve membrane ih štite od djelovanja proteaza.
Opće funkcije bioloških membrana sljedeće:
Oni razgraničavaju sadržaj ćelije od spoljašnje sredine i sadržaj organela od citoplazme.
Oni obezbjeđuju transport tvari u ćeliju i iz nje, od citoplazme do organela i obrnuto.
Oni igraju ulogu receptora (primanje i pretvaranje signala iz okoline, prepoznavanje ćelijskih supstanci, itd.).
Oni su katalizatori (obezbeđuju membranske hemijske procese).
Učestvujte u transformaciji energije.
Opća svojstva bioloških membrana
Bez izuzetka, sve ćelijske membrane su izgrađene prema tome opšti princip: to su tanki lipoproteinski filmovi koji se sastoje od dvostrukog sloja molekula lipida, u koje su uključeni proteinski molekuli. U težinskom smislu, zavisno od vrste membrana, lipidi čine 25-60%, a proteini 40-75%. Mnoge membrane sadrže ugljikohidrate, čija količina može doseći 2-10%.
Biološke membrane vrlo selektivno propuštaju tvari iz okolnog rastvora. Oni prilično lako propuštaju vodu i zarobljavaju većinu tvari topljivih u vodi, a prvenstveno ionizirane tvari ili one koje nose električni naboj. Zbog toga su biomembrane dobri električni izolatori u slanim otopinama.
Membranska podloga je dvostruki lipidni sloj u čijem stvaranju učestvuju fosfolipidi i glikolipidi. Lipidni dvosloj formiraju dva reda lipida, čiji su hidrofobni radikali skriveni unutra, a hidrofilne grupe su okrenute prema van i u kontaktu su sa vodenim medijem. Molekuli proteina su kao da su "otopljeni" u lipidnom dvosloju
Poprečni presjek plazma membrane
Lipidni sastav membrana:
Fosfolipidi. Svi fosfolipidi se mogu podijeliti u 2 grupe - glicerofosfolipidi i sfingofosfolipidi. Glicerofosfolipidi su klasifikovani kao derivati fosfatidne kiseline. Najčešći membranski glicerofosfolipidi su fosfatidilkolini i fosfatidiletanolamini. U membranama eukariotskih ćelija pronađen je ogroman broj različitih fosfolipida, koji su neravnomjerno raspoređeni po različitim ćelijskim membranama. Ova neravnomjernost se odnosi na raspodjelu i polarnih "glava" i acilnih ostataka.
Specifični fosfolipidi unutrašnje membrane mitohondrija su kardiolipini (difosfatidilgliceroli), izgrađeni na bazi glicerola i dva ostatka fosfatidne kiseline. Sintetiziraju ih enzimi unutrašnje membrane mitohondrija i čine oko 22% svih membranskih fosfolipida.
Plazma membrane ćelija sadrže sfingomijeline u značajnim količinama. Sfingomijelini su izgrađeni na bazi ceramida, aciliranog amino alkohola sfingozina. Polarna grupa se sastoji od ostatka fosforne kiseline i holina, etanolamina ili serina. Sfingomijelini su glavni lipidi u mijelinskoj ovojnici nervnih vlakana.
Glikolipidi. Kod glikolipida hidrofobni dio predstavlja ceramid. Hidrofilna grupa - ugljikohidratni ostatak vezan glikozidnom vezom za hidroksilnu grupu na prvom atomu ugljika ceramida. U zavisnosti od dužine i strukture ugljikohidratnog dijela, postoje cerebrozidi, koji sadrže mono- ili oligosaharidni ostatak, i gangliozidi, za čiju OH grupu je vezan kompleksni, razgranati oligosaharid koji sadrži N-acetilneuraminsku kiselinu (NANA).
Polarne "glave" glikosfingolipida nalaze se na vanjskoj površini plazma membrane. Značajne količine glikolipida nalaze se u membranama moždanih stanica, eritrocitima i epitelnim stanicama. Gangliozidi eritrocita različitih pojedinaca razlikuju se po strukturi oligosaharidnih lanaca koji pokazuju antigena svojstva.
Holesterol. Holesterol je prisutan u svim membranama životinjskih stanica. Njegova molekula se sastoji od krutog hidrofobnog jezgra i fleksibilnog ugljikovodičnog lanca, a jedina hidroksilna grupa je "polarna glava".
Za životinjsku ćeliju prosječni molarni omjer holesterol/fosfolipid je 0,3-0,4, ali je u plazma membrani taj odnos mnogo veći (0,8-0,9). Prisustvo holesterola u membranama smanjuje pokretljivost masne kiseline, smanjuje lateralnu difuziju lipida i proteina, te stoga može utjecati na funkcije: membranskih proteina.
U biljnim membranama nema holesterola, ali postoje biljni steroidi - sitosterol i stigmasterol.
Membranski proteini: Uobičajeno je da se dijeli na integralne (transmembranske) i periferne. Integral proteini imaju velike hidrofobne regije na svojoj površini i u njima su netopivi vode. Oni su povezani sa membranskim lipidima hidrofobnim interakcijama i djelomično su uronjeni u debljinu lipidnog dvosloja i često prodiru u dvosloj, ostavljajući ga površine su relativno male hidrofilne površine. Odvojite ove proteine od membrane se mogu postići samo sa deterdžentima kao što su dodecil sulfat ili soli žučne kiseline, koje uništavaju lipidni sloj i pretvaraju protein u rastvorljiv formirati (otopiti ga) formiranjem saradnika s njim. Sve dalje operacije Prečišćavanje integralnih proteina se takođe vrši u prisustvu deterdženata. Periferni proteini su povezani sa površinom lipidnog dvosloja elektrostatičkih sila i može se isprati sa membrane slanim rastvorima.
23. Mehanizmi za prijenos tvari kroz membrane: jednostavna difuzija, pasivni simport i antiport, primarni aktivni transport, sekundarni aktivni transport, regulirani kanali (primjeri). Prenos kroz membranu makromolekula i čestica. Učešće membrana u međućelijskim interakcijama.
Ima ih nekoliko mehanizama transporta supstanci kroz membranu .
Difuzija- prodiranje supstanci kroz membranu duž gradijenta koncentracije (od područja gdje je njihova koncentracija veća do područja gdje je njihova koncentracija niža). Difuzni transport supstanci (voda, joni) odvija se uz učešće membranskih proteina, koji imaju molekularne pore, ili uz učešće lipidne faze (za supstance rastvorljive u mastima).
Sa olakšanom difuzijom specijalni proteini nosači membrane selektivno se vežu za jedan ili drugi ion ili molekul i nose ih preko membrane duž gradijenta koncentracije.
Olakšana difuzija supstanci
Proteini translokaze postoje u ćelijskim membranama. U interakciji sa specifičnim ligandom, osiguravaju njegovu difuziju (transport iz područja veće koncentracije u područje niže koncentracije) preko membrane. Za razliku od proteinskih kanala, translokaze prolaze kroz konformacijske promjene u procesu interakcije s ligandom i njegovom prijenosu kroz membranu. Kinetički, prijenos tvari olakšanom difuzijom sliči enzimska reakcija. Za translokaze postoji koncentracija zasićenja liganda, pri kojoj su sva mjesta vezivanja proteina sa ligandom zauzeta, a proteini rade maksimalnom brzinom Vmax. Stoga, brzina transporta tvari olakšanom difuzijom ne ovisi samo o gradijentu koncentracije transportiranog liganda, već i o broju proteina nosača u membrani.
Postoje translokaze koje prenose samo jednu vodotopivu supstancu s jedne strane membrane na drugu. Takav jednostavan transport se zove "pasivni uniport". Primjer uniporta je funkcioniranje GLUT-1, translokaze koja prenosi glukozu kroz membranu eritrocita:
Olakšana difuzija (uniport) glukoze u eritrocite pomoću GLUT-1 (S - molekula glukoze). Molekul glukoze je vezan nosačem na vanjskoj površini plazma membrane. Dolazi do konformacijske promjene, a centar nosača, okupiran glukozom, biva izložen unutrašnjosti ćelije. Zbog konformacijskih promjena, nosač gubi afinitet za glukozu, a molekul se oslobađa u citosol ćelije. Odvajanje glukoze od nosača uzrokuje konformacijsku promjenu u proteinu, i on se vraća na svoje izvorne "informacije".
Neke translokaze mogu nositi dvije različite tvari duž gradijenta koncentracije u istom smjeru - pasivni simport , ili u suprotnim smjerovima - pasivni antiport .
Primjer translokaze koja djeluje putem pasivnog antiport mehanizma je anionski transporter membrane eritrocita. Unutrašnja mitohondrijalna membrana sadrži mnogo translokaza koje obavljaju pasivni antiport. U procesu takvog prenosa dolazi do ekvivalentne razmene jona, ali ne uvek do ekvivalentne razmene zaduženja.
primarni aktivni transport
Transport nekih neorganskih jona ide protiv gradijenta koncentracije uz učešće transportnih ATPaza (jonske pumpe). Sve jonske pumpe istovremeno služe kao enzimi sposobni za autofosforilaciju i autodefosforilaciju. ATPaze se razlikuju po ionskoj specifičnosti, broju prenesenih jona i smjeru transporta. Kao rezultat funkcioniranja ATPaze, transportirani ioni se akumuliraju na jednoj strani membrane. Ma+,K+-ATPaza, Ca2+-ATPaza i H+,K+,-ATPaza sluznice želuca su najčešće u plazma membrani ljudskih ćelija.
Na+, K+-ATPaza
Ovaj enzim nosilac katalizira ATP-ovisni transport jona Na+ i K+ kroz plazma membranu. Ka+,K+-ATPaza se sastoji od α i β podjedinica; α - katalitička velika podjedinica, i β - mala podjedinica (glikoprotein). Aktivni oblik translokaze je tetramer (αβ)2.
Na+,K+-ATPaza je odgovorna za održavanje visoke koncentracije K+ u ćeliji i niske koncentracije Na+. Budući da Na + D + -ATPaza ispumpava tri pozitivno nabijena jona, a pumpa dva, na membrani nastaje električni potencijal sa negativnom vrijednošću unutar ćelije u odnosu na njenu vanjsku površinu.
Ca2+-ATPaza lokaliziran ne samo u plazma membrani, već iu ER membrani. Enzim se sastoji od deset transmembranskih domena koji pokrivaju ćelijsku membranu. Između druge i treće domene nalazi se nekoliko ostataka asparaginske kiseline koji su uključeni u vezivanje kalcijuma. Područje između četvrte i pete domene ima centar za vezivanje ATP-a i autofosforilaciju na ostatku asparaginske kiseline. Ca2+-ATPaze plazma membrana nekih ćelija su regulisane proteinom kalmodulinom. Svaka od Ca2+-ATPaza plazma membrane i ER je predstavljena sa nekoliko izoforma.
sekundarni aktivni transport
Transport nekih otopljenih tvari protiv gradijenta koncentracije ovisi o istovremenom ili uzastopnom transportu druge tvari duž gradijenta koncentracije u istom smjeru (aktivni simport) ili u suprotnom smjeru (aktivni antiport). U ljudskim ćelijama Na+ je najčešće ion koji se transportuje duž gradijenta koncentracije.

Redoslijed događaja u procesu rada Ca2*-ATP-aze.
1 - vezivanje dva kalcijumova jona na mestu ATP-aze koje je okrenuto prema citosolu;
2 - promjena naboja i konformacije enzima (ATPaze), uzrokovana dodatkom dva Ca2+ jona, dovodi do povećanja afiniteta za ATP i aktivacije autofosforilacije;
3 - autofosforilacija je praćena informacijskim promjenama, ATPaza se zatvara sa unutra membrana i otvara se izvana;
4 - dolazi do smanjenja afiniteta vezivnih centara za jone kalcijuma i oni se odvajaju od ATPaze;
5 - autodefosforilaciju aktiviraju joni magnezijuma, kao rezultat toga, Ca2+-ATP-aza gubi ostatak fosfora i dva Mg2+ jona;
6 - ATPaza se vraća u prvobitno stanje.
Primjer ove vrste transporta je Na+,Ca2+ izmjenjivač plazma membrane (aktivni antiport), natrijum joni se transportuju u ćeliju duž gradijenta koncentracije, a ioni Ca2+ izlaze iz ćelije protiv gradijenta koncentracije.
Prema mehanizmu aktivnog simporta dolazi do apsorpcije glukoze u crijevnim stanicama i reapsorpcije glukoze i aminokiselina iz primarnog urina od strane stanica bubrega.
Transport makromolekula i čestica kroz membranu: endocitoza i egzocitoza
Makromolekule proteina, nukleinskih kiselina, polisaharida, lipoproteinskih kompleksa itd. ne prolaze kroz ćelijske membrane, za razliku od jona i monomera. Transport makromolekula, njihovih kompleksa i čestica u ćeliju odvija se na potpuno drugačiji način - endocitozom. At endocitoza (endo...- iznutra) određeni dio plazmaleme hvata i, takoreći, obavija ekstracelularni materijal, zatvarajući ga u membransku vakuolu koja je nastala kao rezultat invaginacije membrane. Nakon toga, takva vakuola je povezana s lizozomom, čiji enzimi razgrađuju makromolekule do monomera.
Obrnuti proces endocitoze egzocitoza (egzo...- vani). Zahvaljujući njemu, stanica uklanja unutarstanične produkte ili neprobavljene ostatke zatvorene u vakuole ili vezikule. Vezikula se približava citoplazmatskoj membrani, spaja se s njom, a njen sadržaj se oslobađa u okolinu. Kako se izlučuju probavni enzimi, hormoni, hemiceluloza itd.
Dakle, biološke membrane, kao glavni strukturni elementi ćelije, služe ne samo kao fizičke granice, već i kao dinamičke funkcionalne površine. Na membranama organela odvijaju se brojni biohemijski procesi kao što su aktivna apsorpcija supstanci, konverzija energije, sinteza ATP-a itd.
UČEŠĆE MEMBRANA U MEĐUĆELIČNIM INTERAKCIJAMA
Plazma membrana eukariotskih ćelija sadrži mnoge specijalizovane receptore, koji u interakciji sa ligandima izazivaju specifične ćelijske odgovore. Neki receptori vezuju signalne molekule - hormone, neurotransmitere, drugi - nutrijente i metabolite, a treći su uključeni u staničnu adheziju. Ova klasa uključuje receptore neophodne za prepoznavanje i adheziju ćelija, kao i receptore odgovorne za vezivanje ćelija za proteine ekstracelularnog matriksa kao što su fibronektin ili kolagen.
Sposobnost ćelija za specifično međusobno prepoznavanje i adheziju je važna za embrionalni razvoj. Kod odraslih, interakcije adhezivnih stanica-ćelija i stanica-matriks i dalje su bitne za održavanje stabilnosti tkiva. U velikoj porodici receptora stanične adhezije, integrini, selektini i kadherini su najviše proučavani.
Integrins- ekstenzivna superfamilija homolognih receptora na ćelijskoj površini za molekule ekstracelularnog matriksa, kao što su kolagen, fibronektin, laminin, itd. Budući da su transmembranski proteini, oni stupaju u interakciju i sa ekstracelularnim molekulima i sa intracelularnim proteinima citoskeleta. Zbog toga su integrini uključeni u prijenos informacija iz vanćelijske sredine u ćeliju, određujući na taj način smjer njene diferencijacije, oblik, mitotičku aktivnost i sposobnost migracije. Prijenos informacija može ići i u suprotnom smjeru – od intracelularnih proteina preko receptora do ekstracelularnog matriksa.
Primjeri nekih integrina:
receptori za proteine ekstracelularnog matriksa. Vežu se za glikoproteinske komponente ekstracelularnog matriksa, posebno za fibronektin, laminin i vitronektin (vidjeti dio 15);
integrini trombocita (IIb i IIIa) su uključeni u agregaciju trombocita koja se javlja tokom koagulacije krvi;
adhezionih proteina leukocita. Da bi migrirali na mjesto infekcije i upale, leukociti moraju stupiti u interakciju sa vaskularnim endotelnim stanicama. Ova interakcija može posredovati u vezivanju T-limfocita za fibroblaste tokom upale.
Kadherini i selektini porodice transmembranskih Ca 2+-zavisnih glikoproteina uključenih u međućelijsku adheziju. Tri mogući načini učešće receptora ovog tipa u međućelijskoj adheziji.


fibronektinski receptor. Receptor fibronektina pripada porodici integrina. Svaka podjedinica ima jednu transmembransku domenu, kratku citoplazmatsku domenu i proširenu N-ekstracelularnu domenu. Obje podjedinice (α, β) integrina su glikozilirane i držane zajedno nekovalentnim vezama, α-podjedinica se sintetizira kao jedan polipeptidni lanac, koji se zatim cijepa u mali transmembranski lanac i veliki ekstracelularni lanac povezan disulfidom mostova. β-podjedinica sadrži 4 ponavljanja od po 40 aminokiselinskih ostataka. α-podjedinice su bogate cisteinom i sadrže mnoge unutarlančane disulfidne veze (nije prikazano na slici). Vezivanjem za fibronektin izvan i za citoskelet unutar ćelije, integrin djeluje kao transmembranski linker.
Metode interakcije između molekula površine ćelije u procesu međućelijske adhezije. A - receptori jedne ćelije mogu da se vežu za iste receptore susednih ćelija (homofilno vezivanje); B - receptori jedne ćelije mogu da se vežu za receptore druge vrste susednih ćelija (heterofilno vezivanje); B - ćelijski površinski receptori susjednih ćelija mogu komunicirati jedni s drugima koristeći polivalentne molekule povezivača.
Kadherini iz različitih tkiva su vrlo slični, sa 50-60% homolognih aminokiselinskih sekvenci. Svaki receptor ima jednu transmembransku domenu.
Najpotpunije su okarakterisane tri grupe kadherinskih receptora:
E-kadherin se nalazi na površini mnogih ćelija u epitelnim i embrionalnim tkivima;
N-kadherin je lokalizovan na površini nervnih ćelija, srčanih ćelija i sočiva;
P-kadherin se nalazi na ćelijama placente i epiderme.
Kadherini igraju važnu ulogu u početnoj međućelijskoj adheziji, u fazama morfo- i organogeneze, te osiguravaju strukturni integritet i polaritet tkiva, posebno epitelnog monosloja.
U porodici selectin receptorima, tri proteina su najbolje proučavana: L-selektin, P-selektin i E-selektin. Ekstracelularni dio selektina sastoji se od 3 domeni: prvi domen je predstavljen sa 2-9 blokova ponavljajućih aminokiselinskih ostataka (protein regulacije komplementa), drugi je domen epidermalnog faktora rasta (EGF), treći je N-terminalni lektinski domen. Selektini L, P, E se razlikuju po broju blokova u proteinu regulacije komplementa. Lektini su porodica proteina koji specifično stupaju u interakciju s određenim sekvencama ostataka ugljikohidrata u glikoproteinima, proteoglikanima i glikolipidima ekstracelularnog matriksa.
Kratki opis:
Sazonov V.F. 1_1 Struktura stanične membrane [Elektronski izvor] // Kineziolog, 2009-2018: [web stranica]. Datum ažuriranja: 06.02.2018..__.201_). _Opisana je struktura i funkcioniranje ćelijske membrane (sinonimi: plazmalema, plazmolema, biomembrana, ćelijska membrana, vanjska ćelijska membrana, ćelijska membrana, citoplazmatska membrana). Ove početne informacije su neophodne i za citologiju i za razumijevanje procesa nervna aktivnost: nervna ekscitacija, inhibicija, rad sinapsi i senzornih receptora.
ćelijska membrana (plazma A lema ili plazma O lema)
Definicija koncepta
Stanična membrana (sinonimi: plazmalema, plazmolema, citoplazmatska membrana, biomembrana) je trostruka lipoproteinska (tj. "masno-proteinska") membrana koja odvaja ćeliju od okoline i vrši kontroliranu razmjenu i komunikaciju između stanice i okoline.
Glavna stvar u ovoj definiciji nije da membrana odvaja ćeliju od okoline, već samo da ona povezuje ćelije sa okolinom. Membrana je aktivan strukture ćelije, ona stalno radi.
Biološka membrana je ultratanki bimolekularni film fosfolipida obložen proteinima i polisaharidima. Ova ćelijska struktura leži u osnovi barijere, mehaničkih i matriksnih svojstava živog organizma (Antonov VF, 1996).
Figurativni prikaz membrane
Meni stanične membrane Predstavljen je kao rešetkasta ograda sa mnogo vrata u sebi, koja okružuje određenu teritoriju. Bilo koja mala živa bića mogu se slobodno kretati naprijed-nazad kroz ovu ogradu. Ali veći posjetioci mogu ući samo kroz vrata, a ni tada ne svi. Različiti posjetioci imaju ključeve samo od svojih vrata i ne mogu proći kroz tuđa vrata. Dakle, kroz ovu ogradu stalno prolaze tokovi posjetilaca tamo-amo, jer glavna funkcija membrane ograde su dvostruke: da odvoje teritorij od okolnog prostora i istovremeno ga povežu sa okolnim prostorom. Za to ima mnogo rupa i vrata u ogradi - !
Svojstva membrane
1. Propustljivost.
2. Polupropusnost (djelimična propusnost).
3. Selektivna (sinonim: selektivna) propusnost.
4. Aktivna permeabilnost (sinonim: aktivni transport).
5. Kontrolisana propusnost.
Kao što vidite, glavno svojstvo membrane je njena propusnost u odnosu na različite tvari.
6. Fagocitoza i pinocitoza.
7. Egzocitoza.
8. Prisustvo električnih i hemijskih potencijala, tačnije razlike potencijala između unutrašnje i spoljašnje strane membrane. Slikovito se to može reći "membrana pretvara ćeliju u "električnu bateriju" kontrolirajući tokove jona". detalji: .
9. Promjene električnog i hemijskog potencijala.
10. Razdražljivost. Posebni molekularni receptori koji se nalaze na membrani mogu se povezati sa signalnim (kontrolnim) supstancama, zbog čega se može promijeniti stanje membrane i cijele stanice. Molekularni receptori pokreću bio hemijske reakcije kao odgovor na kombinaciju liganada (kontrolnih supstanci) s njima. Važno je napomenuti da signalna supstanca djeluje na receptor izvana, dok se promjene nastavljaju unutar ćelije. Ispostavilo se da je membrana prenosila informacije iz okoline u unutrašnje okruženje ćelije.
11. Katalitička enzimska aktivnost. Enzimi mogu biti ugrađeni u membranu ili povezani sa njenom površinom (unutar i izvan ćelije) i tamo vrše svoju enzimsku aktivnost.
12. Promjena oblika površine i njene površine. To omogućava membrani da formira izrasline prema van ili, obrnuto, invaginacije u ćeliju.
13. Sposobnost stvaranja kontakata sa drugim ćelijskim membranama.
14. Adhezija - sposobnost lijepljenja za čvrste površine.
Kratka lista svojstava membrane
- Propustljivost.
- Endocitoza, egzocitoza, transcitoza.
- Potencijali.
- Razdražljivost.
- enzimska aktivnost.
- Kontakti.
- Adhezija.
Funkcije membrane
1. Nepotpuna izolacija unutrašnjeg sadržaja od spoljašnjeg okruženja.
2. Glavna stvar u radu ćelijske membrane je razmjena razne supstance između ćelije i vanćelijske sredine. To je zbog takve osobine membrane kao što je propusnost. Osim toga, membrana reguliše ovu razmjenu regulacijom njene permeabilnosti.
3. Druga važna funkcija membrane je stvarajući razliku u hemijskim i električnim potencijalima između njegove unutrašnje i vanjske strane. Zbog toga, unutar ćelije ima negativan električni potencijal -.
4. Kroz membranu se također izvodi razmjena informacija između ćelije i njene okoline. Posebni molekularni receptori koji se nalaze na membrani mogu se vezati za kontrolne supstance (hormone, medijatore, modulatore) i pokrenuti biohemijske reakcije u ćeliji, što dovodi do razne promjene u radu ćelije ili u njenim strukturama.
Video:Struktura ćelijske membrane
Video predavanje:Detalji o strukturi membrane i transportu
Struktura membrane
Ćelijska membrana ima univerzalnu troslojni struktura. Njegov srednji masni sloj je kontinuiran, a gornji i donji slojevi proteina ga prekrivaju u obliku mozaika pojedinačnih proteinskih područja. Masni sloj je osnova koja osigurava izolaciju ćelije od okoline, izolujući je od okoline. Sam po sebi, vrlo slabo propušta materije rastvorljive u vodi, ali lako propušta one rastvorljive u mastima. Stoga, propusnost membrane za tvari topljive u vodi (na primjer, ione) mora biti osigurana posebnim proteinskim strukturama - i.
Ispod su mikrofotografije stvarnih ćelijskih membrana kontaktnih ćelija, dobijene pomoću elektronskog mikroskopa, kao i šematski crtež koji prikazuje troslojnu membranu i mozaičnu prirodu njenih proteinskih slojeva. Za uvećanje slike kliknite na nju.

Odvojena slika unutrašnjeg lipidnog (masnog) sloja ćelijske membrane, prožetog integralnim ugrađenim proteinima. Gornji i donji sloj proteina se uklanjaju kako ne bi ometali razmatranje lipidnog dvosloja

Slika iznad: Nepotpuni šematski prikaz ćelijske membrane (ćelijskog zida) sa Wikipedije.
Imajte na umu da su vanjski i unutrašnji slojevi proteina ovdje uklonjeni sa membrane tako da možemo bolje vidjeti središnji masni dvostruki lipidni sloj. U pravoj ćelijskoj membrani, veliki proteinski "otoci" plutaju iznad i ispod masnog filma (male kuglice na slici), a membrana se ispostavi da je deblja, troslojna: proteini-masti-proteini . Tako da je to zapravo kao sendvič od dve proteinske „kriške hleba“ sa debelim slojem „putera“ u sredini, tj. ima troslojnu strukturu, a ne dvoslojnu.
Na ovoj slici male plave i bijele kuglice odgovaraju hidrofilnim (mokrim) "glavama" lipida, a "žice" vezane za njih odgovaraju hidrofobnim (nemočivim) "repovima". Od proteina prikazani su samo integralni membranski proteini od kraja do kraja (crvene globule i žute spirale). Žute ovalne tačke unutar membrane su molekule holesterola. Žuto-zeleni lanci kuglica na vanjskoj strani membrane su oligosaharidni lanci koji formiraju glikokaliks. Glikokaliks je poput ugljikohidratnog ("šećernog") "puha" na membrani, formiranog od dugih ugljikohidratno-proteinskih molekula koji vire iz nje.
Living je mala "proteinsko-masna vrećica" napunjena polutečnim sadržajem nalik na žele, u koju prodiru filmovi i cijevi.
Zidovi ove vrećice formirani su dvostrukim masnim (lipidnim) filmom, prekrivenim iznutra i izvana proteinima - ćelijskom membranom. Stoga se kaže da membrana ima troslojna struktura : proteini-masti-proteini. Unutar ćelije također postoji mnogo sličnih masnih membrana koje dijele njen unutrašnji prostor na odjeljke. Ćelijske organele su okružene istim membranama: jezgrom, mitohondrijama, hloroplastima. Dakle, membrana je univerzalna molekularna struktura svojstvena svim stanicama i svim živim organizmima.

S lijeve strane - više nije pravi, već umjetni model komada biološke membrane: ovo je trenutni snimak dvosloja masnog fosfolipida (tj. dvostrukog sloja) u procesu njegovog molekularno-dinamičkog modeliranja. Prikazana je proračunska ćelija modela - 96 PQ molekula ( f osphatidil X oline) i 2304 molekula vode, ukupno 20544 atoma.
Desno je vizualni model jedne molekule istog lipida, od kojeg je sastavljen membranski lipidni dvosloj. Ima hidrofilnu (vodoljubivu) glavu na vrhu, i dva hidrofobna (vodobojna) repa na dnu. Ovaj lipid ima jednostavno ime: 1-steroil-2-dokozaheksaenoil-Sn-glicero-3-fosfatidilholin (18:0/22:6(n-3)cis PC), ali ga ne morate pamtiti osim ako ne planirajte da vaš učitelj padne u nesvijest dubinom vašeg znanja.
Možete dati precizniju naučnu definiciju ćelije:
je uređen, strukturiran heterogeni sistem biopolimera ograničen aktivnom membranom, koji učestvuje u jednom skupu metaboličkih, energetskih i informacionih procesa, a takođe održava i reprodukuje čitav sistem kao celinu.
Unutar ćelije također prodiru membrane, a između membrana nije voda, već viskozni gel/sol promjenjive gustoće. Zbog toga molekuli u ćeliji ne lebde slobodno, kao u epruveti sa vodeni rastvor, ali uglavnom sjede (imobilizirani) na polimernim strukturama citoskeleta ili intracelularnih membrana. Stoga se kemijske reakcije odvijaju unutar ćelije gotovo kao u čvrstom tijelu, a ne u tekućini. Vanjska membrana koja okružuje ćeliju također je prekrivena enzimima i molekularnim receptorima, što je čini vrlo aktivnim dijelom ćelije.
Ćelijska membrana (plazmalema, plazmolema) je aktivna ljuska koja odvaja ćeliju od okoline i povezuje je sa okolinom. © Sazonov V.F., 2016.
Iz ove definicije membrane slijedi da ona ne ograničava samo ćeliju, već aktivno radi povezujući ga sa svojim okruženjem.
Masnoća koja čini membrane je posebna, pa se njene molekule obično nazivaju ne samo mast, već lipidi, fosfolipidi, sfingolipidi. Membranska folija je dvostruka, odnosno sastoji se od dva filma koja su zalijepljena zajedno. Stoga udžbenici pišu da se baza ćelijske membrane sastoji od dva sloja lipida (ili " dvosloj", tj. dvostruki sloj). Za svaki pojedinačni lipidni sloj, jedna strana se može nakvasiti vodom, a druga ne može. Dakle, ovi filmovi se međusobno lijepe upravo svojim nemokrim stranama.
membrana bakterija
Ljuska prokariotske ćelije gram-negativnih bakterija sastoji se od nekoliko slojeva, prikazanih na donjoj slici.
Slojevi ljuske gram-negativnih bakterija:
1. Unutrašnja troslojna citoplazmatska membrana, koja je u kontaktu sa citoplazmom.
2. Ćelijski zid koji se sastoji od mureina.
3. Spoljna troslojna citoplazmatska membrana, koja ima isti sistem lipida sa proteinskim kompleksima kao i unutrašnja membrana.
Komunikacija gram-negativnih bakterijskih stanica sa vanjskim svijetom kroz tako složenu strukturu u tri koraka ne daje im prednost u preživljavanju u teškim uvjetima u odnosu na gram-pozitivne bakterije koje imaju manje moćnu ljusku. Oni to jednostavno ne prihvataju dobro visoke temperature, povećana kiselost i pad pritiska.




Video predavanje: plazma membrana. E.V. Cheval, Ph.D.
Video predavanje:Membrana kao ćelijska granica. A. Ilyaskin
Važnost membranskih jonskih kanala
Lako je razumjeti da samo tvari topljive u mastima mogu ući u ćeliju kroz membranski masni film. To su masti, alkoholi, gasovi. Na primjer, u eritrocitima kisik i ugljični dioksid lako ulaze i izlaze direktno kroz membranu. Ali voda i tvari topljive u vodi (na primjer, ioni) jednostavno ne mogu proći kroz membranu u bilo koju ćeliju. To znači da su im potrebne posebne rupe. Ali ako samo napravite rupu u masnom filmu, on će se odmah zategnuti. sta da radim? Rješenje je pronađeno u prirodi: potrebno je napraviti posebne strukture za transport proteina i provući ih kroz membranu. Tako se dobijaju kanali za prolaz materija nerastvorljivih u mastima - jonski kanali ćelijske membrane.
Dakle, da dam svoju membranu dodatna svojstva propusnosti za polarne molekule (jone i vodu), ćelija sintetizira posebne proteine u citoplazmi, koji se potom integriraju u membranu. Oni su dva tipa: transporter proteina (na primjer, transport ATPaze) i proteini koji formiraju kanale (tvorci kanala). Ovi proteini su ugrađeni u dvostruki masni sloj membrane i formiraju transportne strukture u obliku transportera ili u obliku jonskih kanala. Kroz ove transportne strukture sada mogu proći različite tvari rastvorljive u vodi, koje inače ne mogu proći kroz film masne membrane.
Općenito se nazivaju i proteini ugrađeni u membranu integral, upravo zato što su, takoreći, uključeni u sastav membrane i prodiru kroz nju. Drugi proteini, koji nisu integralni, formiraju, takoreći, ostrva koja "plutaju" na površini membrane: ili duž njene vanjske površine ili duž njene unutrašnje. Uostalom, svi znaju da je mast dobro mazivo i da se po njoj lako klizi!
zaključci
1. Općenito, membrana je troslojna:
1) spoljni sloj proteinskih "ostrva",
2) masno dvoslojno "more" (lipidni dvosloj), tj. dvostruki lipidni film
3) unutrašnji sloj proteinskih "ostrva".
Ali postoji i labav vanjski sloj - glikokaliks, koji se formira od glikoproteina koji vire iz membrane. Oni su molekularni receptori za koje se vežu signalne kontrole.
2. Posebne proteinske strukture su ugrađene u membranu, osiguravajući njenu propusnost za jone ili druge supstance. Ne smijemo zaboraviti da je more masti na nekim mjestima prožeto integralnim proteinima. A posebno se formiraju integralni proteini transportne strukture ćelijsku membranu (vidi odjeljak 1_2 Mehanizmi membranskog transporta). Preko njih tvari ulaze u ćeliju, a također se iz ćelije uklanjaju prema van.
3. Enzimski proteini mogu se nalaziti na bilo kojoj strani membrane (spoljašnja i unutrašnja), kao i unutar membrane, što utiče kako na stanje same membrane tako i na život cele ćelije.
Dakle, ćelijska membrana je aktivna varijabilna struktura koja aktivno radi u interesu cijele ćelije i povezuje je sa vanjskim svijetom, a nije samo "zaštitna ljuska". Ovo je najvažnija stvar koju treba znati o ćelijskoj membrani.
U medicini se membranski proteini često koriste kao „mete“. lijekovi. Kao takve mete djeluju receptori, jonski kanali, enzimi, transportni sistemi. Nedavno, pored membrane, meta za lekovite supstance takođe postaju geni skriveni u ćelijskom jezgru.
Video:Uvod u biofiziku ćelijske membrane: struktura membrane 1 (Vladimirov Yu.A.)
Video:Povijest, struktura i funkcije ćelijske membrane: Struktura membrana 2 (Vladimirov Yu.A.)
© 2010-2018 Sazonov V.F., © 2010-2016 kineziolog.bodhy.
Main strukturna jedinicaživi organizam - ćelija, koja je diferencirano područje citoplazme, okruženo staničnom membranom. S obzirom na to da ćelija obavlja mnoge važne funkcije, kao što su reprodukcija, prehrana, kretanje, ljuska mora biti plastična i gusta.
Istorija otkrića i istraživanja ćelijske membrane
Godine 1925. Grendel i Gorder su napravili uspješan eksperiment kako bi identificirali "sjene" eritrocita, odnosno prazne ljuske. Uprkos nekoliko učinjenih grubih grešaka, naučnici su otkrili lipidni dvosloj. Njihov rad nastavili su Danielli, Dawson 1935., Robertson 1960. godine. Kao rezultat dugogodišnjeg rada i gomilanja argumenata 1972. godine, Singer i Nicholson su stvorili fluidni mozaični model strukture membrane. Dalji eksperimenti i studije potvrdili su radove naučnika.
Značenje
Šta je ćelijska membrana? Ova riječ se počela koristiti prije više od stotinu godina, a u prijevodu s latinskog znači "film", "koža". Tako označite granicu ćelije, koja je prirodna barijera između unutrašnjeg sadržaja i spoljašnje sredine. Struktura stanične membrane sugerira polupropusnost, zbog koje vlaga i hranjivi sastojci i proizvodi raspadanja mogu slobodno prolaziti kroz nju. Ova ljuska se može nazvati glavnom strukturnom komponentom organizacije ćelije.

Razmotrite glavne funkcije stanične membrane
1. Odvaja unutrašnji sadržaj ćelije i komponente spoljašnjeg okruženja.
2. Pomaže u održavanju konstantnog hemijskog sastava ćelije.
3. Reguliše pravilan metabolizam.
4. Omogućava međusobnu vezu između ćelija.
5. Prepoznaje signale.
6. Funkcija zaštite.
"plazma školjka"
Vanjska ćelijska membrana, također nazvana plazma membrana, je ultramikroskopski film debljine pet do sedam nanometara. Sastoji se uglavnom od proteinskih jedinjenja, fosfolida, vode. Film je elastičan, lako upija vodu, a također brzo vraća svoj integritet nakon oštećenja.
Razlikuje se po univerzalnoj strukturi. Ova membrana zauzima granični položaj, sudjeluje u procesu selektivne propusnosti, izlučivanja produkata raspadanja, sintetizira ih. odnos sa komšijama i pouzdana zaštita unutrašnji sadržaj od oštećenja čini ga važnom komponentom u takvoj stvari kao što je struktura ćelije. Stanična membrana životinjskih organizama ponekad se ispostavi da je prekrivena najtanjim slojem - glikokaliksom, koji uključuje proteine i polisaharide. Biljne ćelije izvan membrane su zaštićene ćelijski zid, koji obavlja funkcije potpore i održavanja oblika. Glavna komponenta njegovog sastava su vlakna (celuloza) - polisaharid koji je nerastvorljiv u vodi.
Dakle, vanjska ćelijska membrana obavlja funkciju popravke, zaštite i interakcije s drugim stanicama.

Struktura ćelijske membrane
Debljina ove pokretne školjke varira od šest do deset nanometara. Stanična membrana ćelije ima poseban sastav, čiju osnovu čini lipidni dvosloj. Hidrofobni repovi, koji su inertni prema vodi, nalaze se iznutra, dok su hidrofilne glave, koje stupaju u interakciju s vodom, okrenute prema van. Svaki lipid je fosfolipid, koji je rezultat interakcije supstanci kao što su glicerol i sfingozin. Lipidna skela je usko okružena proteinima, koji se nalaze u nekontinuiranom sloju. Neki od njih su uronjeni u lipidni sloj, ostali prolaze kroz njega. Kao rezultat, formiraju se vodopropusna područja. Funkcije koje obavljaju ovi proteini su različite. Neki od njih su enzimi, ostali su transportni proteini koji prenose različite tvari iz vanjskog okruženja u citoplazmu i obrnuto.
Ćelijska membrana je prožeta i usko povezana sa integralnim proteinima, dok je veza sa perifernim manje jaka. Ovi proteini obavljaju važnu funkciju, a to je da održavaju strukturu membrane, primaju i pretvaraju signale iz okoline, transportuju supstance i katalizuju reakcije koje se dešavaju na membranama.
Compound
Osnova ćelijske membrane je bimolekularni sloj. Zbog svog kontinuiteta, ćelija ima barijerna i mehanička svojstva. On različite faze ovaj dvosloj može biti poremećen u svojim vitalnim funkcijama. Kao rezultat, formiraju se strukturni defekti kroz hidrofilne pore. U ovom slučaju mogu se promijeniti apsolutno sve funkcije takve komponente kao što je stanična membrana. U tom slučaju, jezgro može patiti od vanjskih utjecaja.

Svojstva
Ćelijska membrana ćelije ima zanimljive karakteristike. Zbog svoje tečnosti, ova ljuska nije kruta struktura, a većina proteina i lipida koji čine njen sastav slobodno se kreće u ravnini membrane.
Generalno, ćelijska membrana je asimetrična, pa je sastav proteinskog i lipidnog sloja različit. Plazma membrane u životinjskim stanicama imaju sloj glikoproteina na svojoj vanjskoj strani, koji obavlja receptorske i signalne funkcije, a također igra važnu ulogu u procesu spajanja stanica u tkivo. Ćelijska membrana je polarna vani naelektrisanje je pozitivno, a iznutra negativno. Pored svega navedenog, ćelijska membrana ima selektivni uvid.

To znači da je osim vode u ćeliju propuštena samo određena grupa molekula i jona otopljenih tvari. Koncentracija tvari kao što je natrij u većini stanica je mnogo niža nego u vanjskom okruženju. Za kalijeve jone karakterističan je drugačiji omjer: njihov broj u ćeliji je mnogo veći nego u okolišu. U tom smislu, joni natrija imaju tendenciju da prodru kroz ćelijsku membranu, a ioni kalija imaju tendenciju da se otpuste van. U takvim okolnostima, membrana aktivira poseban sistem koji ima ulogu "pumpanja", izravnavajući koncentraciju tvari: natrijevi ioni se ispumpavaju na površinu ćelije, a kalijevi ioni se upumpavaju unutra. Ova karakteristika je uključena u najvažnije funkcije stanične membrane.

Ova sklonost jona natrijuma i kalija da se kreću prema unutra s površine igra veliku ulogu u transportu šećera i aminokiselina u ćeliju. U procesu aktivnog uklanjanja iona natrija iz ćelije, membrana stvara uslove za nove prilive glukoze i aminokiselina unutra. Naprotiv, u procesu prenošenja jona kalijuma u ćeliju, broj "transportera" produkata raspadanja iz unutrašnjosti ćelije u spoljašnju sredinu se popunjava.
Kako se ćelija hrani kroz ćelijsku membranu?
Mnoge ćelije uzimaju supstance kroz procese kao što su fagocitoza i pinocitoza. U prvoj varijanti fleksibilna vanjska membrana stvara malo udubljenje u kojem se nalazi zarobljena čestica. Tada promjer udubljenja postaje sve veći sve dok okružena čestica ne uđe u ćelijsku citoplazmu. Fagocitozom se hrane neke protozoe, poput amebe, kao i krvne ćelije- leukociti i fagociti. Slično, ćelije apsorbiraju tekućinu koja sadrži potrebne hranjive tvari. Ovaj fenomen se naziva pinocitoza.
Vanjska membrana je usko povezana sa endoplazmatskim retikulumom ćelije.
Kod mnogih tipova osnovnih komponenti tkiva, izbočine, nabori i mikrovili nalaze se na površini membrane. Biljne ćelije sa vanjske strane ove ljuske prekrivene su drugom, debelom i jasno vidljivom pod mikroskopom. Vlakna od kojih su napravljena pomažu u održavanju tkiva. biljnog porijekla, na primjer, drvo. Životinjske ćelije također imaju brojne vanjske strukture koje se nalaze na vrhu ćelijske membrane. Isključivo su zaštitne prirode, primjer za to je hitin sadržan u integumentarnim stanicama insekata.
Pored ćelijske membrane postoji i unutarćelijska membrana. Njegova funkcija je da podijeli ćeliju u nekoliko specijaliziranih zatvorenih odjeljaka – odjeljaka ili organela, gdje se mora održavati određena sredina.

Stoga je nemoguće precijeniti ulogu takve komponente osnovne jedinice živog organizma kao što je ćelijska membrana. Struktura i funkcije podrazumijevaju značajno proširenje ukupne površine ćelije, poboljšanje metaboličkih procesa. Ova molekularna struktura sastoji se od proteina i lipida. Odvajajući ćeliju od vanjskog okruženja, membrana osigurava njen integritet. Uz njegovu pomoć, međućelijske veze se održavaju na dovoljno jakom nivou, formirajući tkiva. S tim u vezi, možemo zaključiti da jednu od najvažnijih uloga u ćeliji ima ćelijska membrana. Struktura i funkcije koje obavlja radikalno se razlikuju u različitim ćelijama, ovisno o njihovoj namjeni. Kroz ove karakteristike postiže se raznovrsnost fiziološke aktivnosti ćelijskih membrana i njihove uloge u postojanju ćelija i tkiva.