Физиологические функции организма человека. Физические и физиологические свойства скелетных, сердечной и гладких мышц. Учебное пособие «общая физиология»
Лекция 1
Введение в курс
Предмет и задачи физиологии
целенаправленно создавать новые лекарственные препараты . Поэтому студентам, изучающим фармацевтическую химию, необходимо знать современные представления о механизмах фармакологического и токсического воздействия на организм и уметь применять полученные знания в будущей производственной деятельности.
Основные физиологические понятия
Каждая научная дисциплина имеет собственный понятийный аппарат. Естественно, что и в физиологии, науке сформировавшейся еще в XIX веке, существуют собственные понятия и термины. Ниже приведены наиболее общие из них. Другие, термины и понятия, имеющие более узкое значение, будут рассмотрены в ходе изложения соответствующих разделов физиологии на последующих лекциях.
Основные физиологические понятия.
Функция - специфическая деятельность системы или органа. Например, функциями желудочно-кишечного тракта являются моторная, секреторная, всасывательная; функцией системы дыхания - обмен СО 2 и кислорода; функцией системы кровообращения - движение крови по сосудам; функцией миокарда - сокращение и расслабление, и т.д.
Процесс - совокупность последовательных действий или состояний, направленных на достижение определенного результата.
Механизм - способ регулирования процесса или функции. ‚ физиологии принято выделять нервный и гуморальный (т.е. посредством выделения гормонов или других химических биологически активных веществ) механизмы регуляции.
Регуляция - изменение интенсивности и направленности функции (процесса) с целью обеспечения оптимальной деятельности органов и систем.
Система в физиологии это совокупность органов, тканей или других структурных образований, связанных общей функцией. Например, сердечно-сосудистая система обеспечивает с помощью сердца и сосудов доставку тканям питательных, регуляторных, защитных веществ и кислорода, а также отвод продуктов обмена; экспортная (секреторная) система клетки обеспечивает с помощью эндоплазматического ретикулума и аппарата Гольджи синтез и транспортировку из клеток гормонов, липопротеидов и других секретов.
Секрет - специфический продукт жизнедеятельности клетки, выполняющий определенную функцию и выделяющийся во внутреннюю среду организма. Процесс выработки и выделения секрета называется секрецией . По составу секрет делят на белковый (серозный), слизистый (мукоидный), смешанный и липидный.
Раздражитель - факторы внешней и внутренней среды или их изменение, которые оказывают на органы и ткани влияния, приводящие к изменению активности последних. Различают раздражители механические, электрические, химические, температурные, звуковые и т.д. Раздражитель может быть по величине пороговым , т.е. оказывать минимальное эффективное воздействие; максимальным , дальнейшее увеличение силы которого не приводит к увеличению соответствующей реакции; сверхсильным , действие которого может оказывать повреждающий или болевой эффект или приводить к неадекватным ощущениям.
Раздражение - воздействие на живую ткань внешних и внутренних раздражителей.
Реакция - изменение (усиление или ослабление) деятельности организма или его составляющих в ответ на раздражение.
Рефлекторная реакция - ответное действие (или процесс) в организме (системе, органе, ткани, клетке), вызванное рефлексом.
Рефлекс - возникновение или изменение функциональной активности органов, тканей или целостного организма, осуществляемое при участии центральной нервной системы в ответ на раздражение нервных окончаний (рецепторов).
Возбудимость - способность живых клеток воспринимать изменения внешней среды и отвечать на эти изменения реакцией возбуждения. Чем ниже пороговая сила раздражителя, тем выше возбудимость, и наоборот.
Возбуждение - активный физиологический процесс, которым некоторые живые клетки (нервные, мышечные, железистые) отвечают на внешнее воздействие.
Возбудимые ткани - ткани, способные в ответ на внешнее воздействие, превышающее порог возбуждения , переходить из состояния физиологического покоя в состояние возбуждения. ‚ принципе, все живые клетки обладают той или иной степенью возбудимости, но в физиологии к этим тканям принято относить нервную, мышечную, железистую.
Физиология клетки
Клеточные мембраны.
Лекция 2
Трансмембранный транспорт.
Разность электрических потенциалов (в вольтах или мв) между жидкостью, находящейся по одну сторону мембраны и жидкостью по другую ее сторону называется мембранным потенциалом (МП) и обозначается Vм . Величина МП живых клеток составляет обычно от -30 до -100 мв и вся эта разность потенциалов создается в областях непосредственно прилегающих с обоих сторон к клеточной мембране. Уменьшение величины МП называют деполяризацией , увеличение - гиперполяризацией , восстановление исходного значения после деполяризации - реполяризация . Мембранный потенциал существует во всех клетках, но в возбудимых тканях (нервных, мышечных, железистых), мембранный потенциал или как его еще называют в этих тканях, мембранный потенциал покоя , играет ключевую роль в реализации их физиологических функций. Мембранный потенциал обусловлен двумя основными свойствами всех эукариотических клеток: 1) асимметричным распределением ионов между вне- и внутриклеточной жидкостью, поддерживаемым метаболическими процессами; 2) Избирательной проницаемостью ионных каналов клеточных мембран. Чтобы уяснить себе как возникает МП представим себе, что некий сосуд разделен на два отсека мембраной, проницаемой только для ионов калия. Пусть в первом отсеке содержится 0,1 М, а во втором 0,01 М раствор КСl. Поскольку концентрация ионов калия (К +) в первом отсеке в 10 раз выше, чем во втором, то в начальный момент на каждые 10 ионов К + диффундирующих из отсека 1 во второй будет приходится один ион диффундирующий в обратном направлении. Так как анионы хлора (Сl-) не могут переходить через мембрану вместе с катионами калия, то во втором отсеке будет образовываться избыток положительно заряженных ионов и, напротив, в отсеке 1 окажется избыток ионов Сl-. В результате возникает трансмембранная разность потенциалов , препятствующая дальнейшей диффузии К + во второй отсек, поскольку для этого им нужно преодолеть притяжение отрицательных ионов Сl-, в момент вхождения в мембрану со стороны отсека 1 и отталкивание одноименных ионов на выходе из мембраны в отсек 2. Таким образом, на каждый ион К + , проходящий через мембрану в этот момент действуют две силы - химический градиент концентраций (или химическая разность потенциалов), способствующая переходу ионов калия из первого отсека во второй, и электрическая разность потенциалов, заставляющая ионы К + двигаться в обратном направлении. После того как эти две силы уравновесятся, количество ионов К + перемещающееся из отсека 1 в отсек 2 и обратно сравняется, установится электрохимическое равновесие . Соответствующая такому состоянию трансмембранная разность потенциалов называется равновесным потенциалом , в данном конкретном случае равновесным потенциалом для ионов калия (Ек ). В конце 19 века Вальтер Нернст установил, что равновесный потенциал зависит от абсолютной температуры, валентности диффундирующего иона и от отношения концентраций данного иона по разные стороны мембраны:
где Ех- равновесный потенциал для иона X, R - универсальная газовая постоянная = 1,987 кал/(моль град), T - абсолютная температура в градусах Кельвина, F - число Фарадея = 23060 кал/в, Z - заряд переносимого иона, [X] 1 и [X] 2 - концентрации иона в отсеках 1 и 2.
Если перейти от натурального логарифма к десятичному, то для температуры 18˚С и моновалентного иона можно записать уравнение Нернста следующим образом:
Ех= 0,058 lg
Рассчитаем с помощью уравнения Нернста калиевый равновесный потенциал для воображаемой клетки, приняв, что внеклеточная концентрация калия [К + ]н= 0,01 М, а внутриклеточная - [К + ]в = 0,1 М:
Ек= 0,058 lg = 0,058 lg =0,058 (-1) = -0,058 ‚= -58 мв
В данном случае, Ек отрицателен, поскольку ионы калия будут выходить из гипотетичной клетки, заряжая отрицательно слой цитоплазмы, прилегающий к внутренней стороне мембраны. Поскольку в данной гипотетичной системе имеется только один диффундирующий ион, то калиевый равновесный потенциал будет равен мембранному потенциалу (Ек= Vм ).
Приведенный механизм ответственен и за образование мембранного потенциала в реальных клетках, но в отличие от рассмотренной упрощенной системы, в которой через "идеальную" мембрану мог диффундировать только один ион, реальные клеточные мембраны пропускают в той или иной все неорганические ионы. Однако, чем менее мембрана проницаема для какого-либо иона, тем меньшее влияние он оказывает на МП. Учитывая это обстоятельство, Голдманом в 1943г. было предложено уравнение для расчета величины МП реальных клеток, учитывающее концентрации и относительную проницаемость через плазматическую мембрану всех диффундирующих ионов:
Vм = 0,058 lg
Используя метод меченых изотопов, Ричард Кейнс в 1954 г. определил проницаемость клеток мышц лягушки для основных ионов. Оказалось, что проницаемость для натрия примерно в 100 раз меньше, чем для калия, а ион Сl-не вносит заметного вклада в создание МП мышечных клеток. Поэтому для мембран мышечных клеток уравнение Голдмана можно записать в следующем упрощенном виде:
Vм = 0,058 lg
Vм = 0,058 lg
Исследования с применением вводимых в клетки микроэлектродов, показали, что потенциал покоя клеток скелетных мышц лягушки колеблется от -90 до -100 мв. Такое хорошее соответствие экспериментальных данных теоретическим подтверждает, что потенциал покоя определяется диффузионными потоками неорганических ионов. При этом, в реальных клетках мембранный потенциал близок к равновесному потенциалу иона, который характеризуется максимальной трансмембранной проницаемостью, а именно к равновесному потенциалу иона калия.
Роль кальция
Лекция 3
Основы биоэнергетики.
Химическая энергия, особенности ее образования и утилизации в живых системах.
Молекулярные механизмы преобразования энергии в живых клетках изучает биоэнергетика. Восстановленные органические соединения, служащие источником энергии живым организмам, окисляются до СО 2 и Н 2 О, а высвобождающаяся энергия используется для синтеза АТФ и выполнения различных работ.
Прежде чем перейти к рассмотрению конкретных биоэнергетических процессов в клетке вспомним, что в соответствии со вторым законом термодинамики энтропия любой замкнутой системы с течением времени может только возрастать. Энтропия - термодинамическая категория, характеризующая степень неупорядоченности системы. Поэтому, возрастание энтропии, по сути, есть увеличение степени неупорядоченности системы, т.е. переход от более упорядоченной структуры к менее упорядоченной и снижение доли полезной энергии (т.е. энергии, с помощью которой может быть совершена работа). Другими словами из второго закона термодинамики следует, что в любой изолированной системе степень неупорядоченности может только возрастать, а величина полезной (свободной) энергии - уменьшаться . В тоже время живые организмы, по мере того как проходят развитие от оплодотворенной яйцеклетки до состояния взрослой особи, постоянно усложняют свою структуру. Казалось бы, в живых системах второй закон термодинамики не выполняется. Однако, следует иметь ввиду, что этот закон относится к замкнутой системе, а живой организм является открытой системой, для которых характерен постоянный обмен с окружающей средой веществом и энергией и поддерживание относительно низкого уровня энтропии, за счет повышения ее в окружающей среде. Способность извлекать полезную энергию из окружающей среды является одним из тех замечательных свойств, которые отличают живые организмы от неживой природы. Потребляя растительную или животную пищу, живой организм превращает содержащиеся в ней высокоорганизованные молекулы в СО 2 , Н 2 О и низкомолекулярные азотсодержащие соединения, имеющие значительно более высокий уровень энтропии, а образующуюся при этом энергию использует, в том числе на развитие и рост собственного организма, сопровождающийся понижением энтропии. Первичным же источником энергии на земле для большинства организмов является солнце, энтропия которого постоянно повышается и только незначительная часть выделяемой при этом энергии улавливается биосферой и утилизируется растениями и некоторыми бактериями в реакциях фотосинтеза, а остальная безвозвратно теряется во Вселенной. Даже открытая недавно обильная жизнь глубоко на дне океанов в области океанических трещин на т.н. «черных курильщиках» зависит от солнца, поскольку является аэробной, а кислород берется в атмосфере и океане только благодаря процессу фотосинтеза.
В процессе эволюции в организме животных и естественно человека сформировалось множество путей получения, транспорта и утилизации энергии, и в подавляющем большинстве из них для аккумуляции химической энергии используются специальные соединения – фосфагены , имеющие в своей структуре так называемую высокоэнергетическую или макроэргическую связь. Аденозин-5"-трифосфат (АТФ ) представляет собой наиболее распространенный и важнейший представитель этой группы соединений. Кроме АТФ, высокоэнергетические связи имеют ряд других органических полифосфатов, например ГТФ, ЦТФ Для обозначения макроэргической связи используется символ ~. Так молекулу АТФ сокращенно обозначают А-Р-Ф~Ф~Ф (А -аденин, Р -рибоза). Некоторые бактерии даже запасают энергию в форме неорганического полиметафосфата – линейного полимера неопределенной длинны.
При рН=7,0 АТФ представляет анион с высоким зарядом. Трифосфатная группа содержит 4 ОН группы способные к ионизации. 3 из них имеют низкое значение рК и при рН=7 полностью диссоциируют, четвертая группа диссоциирует на 75%. Высокая концентрация отрицательных зарядов является важным фактором, обуславливающим высокоэнергетическую природу АТФ. Эти заряды располагаются близко друг к другу и между ними существует сильное отталкивание. При гидролитическом отщеплении концевой фосфатной группы силы отталкивания ослабевают. Образовавшиеся продукты не стремятся воссоединиться. Вторая важная особенность, обуславливающая большую отрицательную величину ΔG" гидролиза АТФ заключается в том, что оба продукта гиролиза стабилизированы за счет сопряжения. Впрочем, в клетках АТФ почти не встречается в форме свободных ионов, а чаще в виде комплекса с ионами Mg или Mn.
Понятие энергии фосфатной связи, которым пользуются биохимики не следует путать с понятием энергии связи, под которой в физической химии понимают энергию необходимую для разрыва в молекуле связи между 2 атомами. Под энергией макроэргической фосфатной связи понимают разность между свободной энергией исходных веществ и свободной энергией продуктов реакции, в процессе которой фосфорилированное соединение гидролизуется с образованием неорганического фосфата.
Перенос концевой фосфатной группы АТФ на воду сопровождается освобождением свободной энергии , примерно вдвое большей, чем при гидролизе обычной фосфоэфирной связи, например в аденозинмонофосфате. Свободная энергия характеризует ту долю полной энергии заключенной в веществе, с помощью которой может быть совершена работа в изотермических и изобарических условиях , т.е. при постоянной температуре и давлении, а именно такие условия характерны для биологических систем. Изменение свободной энергии измеряется в ккал/моль и обозначается ΔG" если процесс протекает в стандартных условиях (рН=7, Т=25 ºС, субстраты и продукты реакции взяты в одинаковой концентрации равной 1 моль/л) и символом ΔG для реакции в конкретных условиях реальных биологических систем. В случае гидролиза АТФ, ΔG" = -7 ккал/моль, а ΔG = -11-13 ккал/моль Живые организмы используют энергию АТФ для выполнения механической работы мышц, создания концентрационных градиентов и трансмембранных электрических потенциалов.
Конкретные механизмы, обеспечивающие протекание данных процессов будут изложены в последующих лекциях, а теперь рассмотрим, общие принципы, обеспечивающие возможность использования энергии АТФ для совершения химической работы. Представим себе типичный биосинтетический, т.е. анаболический процесс (анаболизм от греч. подъем те процессы синтеза, построения структурных частей клетки, противоположен катаболизму), при котором два мономера -Х и Y- должны соединиться друг с другом в реакции дегидратации (называемой также конденсацией), сопровождающейся выделением воды:
Х-ОН + Y-Н X-Y + Н 2 О + ΔG (3 ккал/моль)
Большинство таких реакций являются эндергоническими (не путать с эндотермическими) и сопровождаются возрастанием свободной энергии на величинуΔG , а движущей силой любой химической реакции является уменьшение свободной энергии и самопроизвольно протекают только экзергонические реакции и, следовательно, почти всегда предпочтительнее обратная катаболическая реакция (называемая гидролизом), поскольку в этом случае имеет место выделение свободной энергии равное по абсолютной величине ее потреблению в реакции конденсации. Поэтому в клетке такие реакции протекают в несколько стадий, в результате которых происходит сопряжение энергетически невыгодного синтеза с сильно экзергоническиой реакцией, протекающей с участием АТФ. Механизм сопряжения реализуется следующим образом. Реакции синтеза, приведенной выше и сопровождающейся возрастанием свободной энергии (предположим ΔG" = 3 ккал/моль ), предшествует фосфорилирование одного из субстратов молекулой АТФ .
А-Р-Ф~Ф~Ф + ХОН → А-Р-Ф~Ф + Х-О-Ф - ΔG = 4 ккал/моль
ΔG" этой реакции значительно меньше (4 ккал/моль ), чем реакции гидролиза АТФ и переноса фосфатной группы на воду (примерно 11 ккал/моль ). Остальная энергия (7 ккал/моль ) аккумулируется в молекуле активированного субстрата, который вовлекается в последующую синтетическую реакцию. Однако, теперь свободная энергия исходных веществ выше, чем свободная энергия продуктов реакции (7 - 3 = 4 ккал/моль) и, реакция протекает слева направо, т.е. в направлении синтеза, сопровождаясь уменьшением свободной энергии:
Х-О-Ф + Y-Н → X-Y + Ф н - ΔG = (4 ккал/моль)
Таким образом, за счет накопленной в молекуле АТФ энергии эндергоническая реакция замещается двумя сопряженными экзергоническими реакциями.
Другой путь использования АТФ для совершения химической работы реализуется через последовательно сопряженные или как их называют тандемные реакции. В этом случае эндергоническая реакция протекает благодаря следующей за ней экзергонической реакции. Возможность протекания тандемных реакций обусловлена тем обстоятельством, что увеличение свободной энергии не исключает протекание химической реакции, а лишь указывает на то, что величина константы равновесия такой реакции меньше единицы, т.е. если имеет место реакция:
А + B C + D + ΔG" 1 (1)
то К eq = < 1
и, следовательно, равновесие реакции смещено влево и это смещение тем более выражено, чем больше по абсолютной величине ΔG" . Однако если за реакцией 1 следует реакция 2
С +X CX- ΔG" 2 (2)
и для этой реакции величина ΔG" сильно отрицательна, то промежуточный продукт реакции С будет потребляться, смещая при этом равновесие реакции 1 вправо. В том случае если суммарная свободная энергия двух реакций будет величиной отрицательной они будут протекать в направлении потребления субстратов А и В и образования продукта СХ.
Теперь рассмотрим где в клетке локализованы механизмы получения энергии и перевода фосфагенов из "незаряженной" формы (например АДФ) в "заряженную" (АТФ) и как они функционируют.
Цикл трикарбоновых кислот
В большинстве клеток и тканей аэробных организмов, образующаяся в результате гликолиза пировиноградная кислота поступает в матрикс митохондрий , где она декарбоксилируется, т.е. от нее отщепляется молекула углекислого газа, а оставшаяся двухуглеродная ацетильная группа соединяется с коферментом А (СоА ), образуя ацетил-СоА . Энергия, выделяющаяся в результате этой реакции, расходуется на перевод специальной молекулы - переносчика водорода из окисленной в восстановленную форму. Переносчик водорода называется никотинамидадениндинуклеотид и обозначается сокращенно NAD (НАД ) или NAD+(окисленная форма) и NADH (НАДН ) или NADH + H+(восстановленная форма). Таким образом, молекулы NAD+ и сходного с ним по структуре и функции флавинадениндинуклеотида , сокращенно FAD , подобно фосфагенам способны запасать и переносить энергию, но не в виде макроэргической фосфатной связи, а в виде активированных атомов водорода. Образовавшийся из пировиноградной кислоты ацетил-СоА подвергается дальнейшему окислению в цикле трикарбоновых кислот . В результате первой реакции цикла (Рис.) из ацетил-С и щавелевоуксусной кислоты (в водной среде существует в виде иона - оксалоацетата) содержащей 4 атома углерода при участии фермента цитратсинтазы образуется трикарбоновая, шестиуглеродная лимонная кислота (цитрат). Поэтому цикл трикарбоновых кислот называют иногда цикл лимонной кислоты. Затем лимонная кислота превращается в изолимонную (изоцитрат). В результате следующей, реакции декарбоксилирования шестиуглеродная молекула изолимонной кислоты превращается в пятиуглеродную α-оксоглутаровую кислоту (α-оксоглутарат ) и происходит восстановление NAD+ в NADH. Далее α-оксоглутаровая кислота декарбоксилируется до янтарной (сукцинат). При этом выделяющаяся энергия не рассеивается в виде тепла, а затрачивается на восстановление еще одной молекулы NAD+ в NADH и образование промежуточного продукта с высокоэнергетической фосфатной связью, который обеспечивает фосфорилирование гуанозиндифосфата (GDP)в гуанозинтрифосфат (GTP).
В результате трех следующих реакций янтарная кислота последовательно превращается в фумаровую кислоту (фумарат ), яблочную кислоту (малат ) и щавелевоуксусную и восстановливается по молекуле FAD и NAD.
С точки зрения биоэнергетики при каждом обороте цикла удаляются 2 атома углерода вошедшего в цикл как ацетил-СоА в виде СО 2 и генерируется 8[H] + 2[H] на входе – это активированный водород -источник энергии.
Таким образом, ферментативные реакции цикла трикарбоновых кислот протекают так, что субстрат первой реакции - щавелевоуксусная кислота является и продуктом последней восьмой реакции, т.е. регенерируется после каждого оборота цикла и процесс запускается вновь. В результате элементарного цикла два углеродных атома, имевшихся в ацетил-СоА, превращаются в СО 2 . Циклический характер последовательности реакции окисления ацетил-СоА и ее основные этапы установил в 1937 году английский биохимик немецкого происхождения, лауреат Нобелевский премии (1953) Ханс Кребс. Поэтому процесс окисления ацетил-СоА имеет еще одно название - цикл Кребса . Благодаря тому, что окисление совершается постепенно, через серию относительно небольших изменений свободной энергии, выделяющаяся энергия не рассеивается в виде бесполезного тепла, а затрачивается на восстановление трех молекул NAD в NADH и одной молекулы FAD в FADH и образование высокоэнергетической фосфатной связи. Эта связь образуется в результате фосфорилирования гуанозиндифосфата в гуанозинтрифосфат. Последний легко обменивается фосфатной группой с АДФ с образованием АТФ.
Следует иметь в виду, что источником ацетил-CoA для окисления в ЦТК, наряду с гликолизом являются и другие биохимические процессы протекающие в клетках, в частности окисление аминокислот и жирных кислот (а также этанол). Четыре молекулы NADH, включая и образовавшуюся при окислении пирувата в ацетил-СоА, и молекула FADH, поступают в дыхательную цепь, где переносимые ими атомы водорода окисляются молекулярным кислородом до воды. Выделяемая при этом энергия затрачивается на синтез АТФ из АДФ и неорганического фосфата в результате сложнейшего, во многом неясного процесса, получившего название окислительного фосфорилирования .
Лекция 4
Лекция 5.
Лекция 6.
Лизосомы: структура и пути образования в клетке, классификация.
Лизосомы представляют собой гетерогенную (разнородную) группу цитоплазматических вакуолеподобных структур размером 1-3 мкм, отличительной особенностью которых является наличие в них кислой среды и большого количества различных гидролаз - ферментов способных расщеплять (гидролизовать) основные типы макромолекул. Присутствие в лизосомах гидролаз определяет их основную функцию в клетке - расщепление макромолекул и более крупных образований как поступающих в клетку из внеклеточного пространства так и имеющих внутриклеточное происхождение. Интересно, что у растений нет лизосом.
Известны три возможных пути образования лизосом в клетке. В каждом случае образуются морфологически различные образования, расщепляющие материал из различных источников.
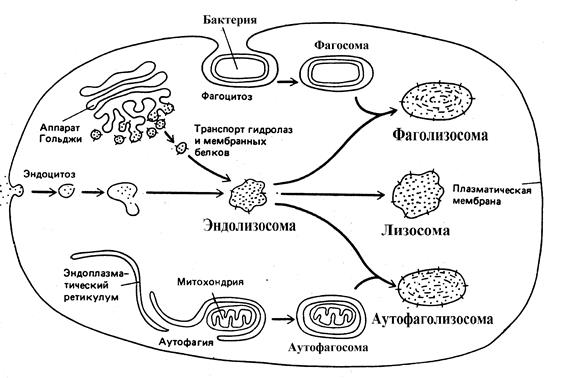 Рис. 6 Пути образования лизосом в клетке
Рис. 6 Пути образования лизосом в клетке
В первом случае расщепляемый материал - белки, полинуклеотиды или полисахариды - попадает в клетку путем эндоцитоза . В ходе этого процесса молекулы, имеющие достаточно крупные размеры и неспособные проникать через мембраны, постепенно окружаются небольшим участком плазмалеммы, который сначала впячивается (инвагинируется), а затем отщепляется вовнутрь клетки, образуя пузырек, содержащий захваченный клеткой материал. Пузырьки образующиеся в результате эндоцитоза получили название эндосомы . По мере движения эндосомы от клеточной мембраны во внутрь клетки она многократно взаимодействует с транспортными пузырьками доставляющими от транс -поверхности аппарата Гольджи гидролитические ферменты и мембранные белки, превращаясь в эндолизосому . Процесс образования и трансформации эндосомы длится около 15 мин и сопровождается закислением внутренней среды, благодаря закачиванию ионов Н+из цитозоля во внутрь эндосомы АТФ-зависимым протонным насосом, функционирующим подобно АТФ -азе внутренней мембраны митохондрий.
Второй путь формирования лизосом называется аутофагией . В процессе аутофагии происходит разрушение отработанных частей самой клетки. Известно, например, что в клетках печени среднее время жизни одной митохондрии составляет около 10 дней, после чего она должна быть утилизирована в лизосомах. На электронных микрофотографиях нормальных клеток можно увидеть лизосомы содержащие митохондрии на разных стадиях деградации. Выше уже отмечалось, что путем аутофагии из клеток печени удаляется избыток гладкого ЭР, после прекращения поступления и выведения из организма ксенобиотиков - индукторов. Процесс аутофагии, по-видимому, начинается с окружения органеллы мембранами, поставляемыми из ЭР, в результате чего образуется аутофагосома . Затем, полагают, что аутофагосома сливается с эндолизосомой, образуя аутофаголизосому, в которой и происходит процесс деградации фрагмента ЭР или другой органеллы.
Третий путь формирования лизосом имеется только у клеток, специализированных для фагоцитоза больших частиц и микроорганизмов. Такие клетки-фагоциты, а к ним относятся клетки крови - нейтрофилы и моноциты, могут поглощать из внеклеточного пространства крупные объекты, образуя фагосомы . Далее фагосома превращается в фаголизосому тем же путем, что и аутофагосома, т.е. сливаясь с эндолизосомой.
Эндосомы, аутофагосомы и фагосомы часто называют общим термином - прелизосомы , а эндолизосомы, аутофаголизосомы и фаголизосомы термином лизосомы. В зрелых лизосомах происходит деградация поглощенного материала до отдельных молекул, например аминокислот, которые поступают в цитозоль и вовлекаются в последующие биохимические превращения. Фрагменты собственной плазматической мембраны не подвергаются воздействию гидролаз и возвращается обратно в плазмалемму с помощью транспортных пузырьков, еще до окончательного формирования лизосомы. Неперевариваемые продукты остаются и накапливаются в лизосомах, которые теряют гидролитические ферменты и превращаются в постлизосомы или остаточные тельца . С возрастом, в клетках человека и животных увеличивается количество остаточных телец, содержащих большое количество липофусцина или пигмента старения.
Липофусцинпредставляет собой биополимеры различной природы, неподдающиеся дальнейшему расщеплению поскольку химические связи между отдельными мономерами образовались не в нормальных биохимических реакциях, а в результате спонтанных окислительных процессов, главным образом свободнорадикальных. Различные заболевания, воздействие радиации и других негативных факторов внешней среды ускоряют процесс накопления пигмента старения.
Клеточное ядро и геном
Лекция 7
Лекция 1
Введение в курс
Предмет и задачи физиологии
Физиология (от греческих слов: физис - природа, логос - учение, наука) наука о функциях как организма в целом, так и отдельных составляющих его систем: органов, тканей, клеток; процессах и механизмах, обуславливающих возможность реализации и регуляции этих функций и обеспечивающих жизнедеятельность человека и животных в их взаимодействии с окружающей средой. Задача физиологии заключается в установлении физических и химических основ процессов жизнедеятельности на всех уровнях организации живых систем от субклеточного до уровня целого организма. О связи со сложностью предмета изучения физиология подразделяется на несколько разделов. К ним относятся: общая и клеточная физиология, задачей которой является изучение закономерностей основных жизненных процессов и механизмов характерных для всех функциональных и структурных систем организма и частная физиология, в свою очередь подразделяющаяся на физиологию отдельных органов: сердца, почек, печени и т.д.; физиологию функциональных систем: дыхания, кровообращения, пищеварения, эндокринной системы, физиологию развития, нейрофизиологию и физиологию поведения, сенсорную физиологию. Существует также прикладная физиология: спорта, труда, питания, космическая и подводная физиология.
Каково же практическое значение физиологии? Почему так важно знать, как работает организм? Наиболее очевидная причина заключается в том, что физиология, и особенно учение о функционировании организма человека, служит научной основой медицины и фармацевтической химии. На всех прежних этапах развития цивилизации подход к лечению болезней был почти целиком эмпирический, т.е. сводился к методу проб и ошибок. В результате, постепенно было подмечено, что при тех или иных болезнях помогают определенные средства - будь то химические или растительные препараты, минеральная вода и горячие ванны, акупунктура или даже методы психологического воздействия, которые широко использовались в народной медицине. Лечебный эффект многих современных препаратов был сначала обнаружен чисто эмпирически (например, нитроглицерин, использовался Нобелем для производства динамита, при этом вызывал у рабочих сильное падение артериального давления, часто приводившее к летальному исходу), а главные механизмы, лежащие в основе целебного действия, целого ряда из них пока так и остаются неизвестными. Однако по мере развития и углубления нашего понимания механизмов функционирования и патологии органов и тканей, снижается роль чисто эмпирических находок и появляется возможность разрабатывать эффективные, научно обоснованные способы лечения заболеваний человека, целенаправленно создав
1.1. Общее представление о физиологии
Основные вопросы : Организм и среда обитания. Поняте о физиологии. Физиологическая функция и ее мультипараметрическая характеристика. Гомеостаз, релаксоконстанты и их виды. Методы исследования в физиологии. Острый и хронический эксперименты. Аналитический и системный подходы к изучению физиологических функций. Теория функциональных систем по П.К. Анохину.
Организм человека - целостная, саморегулирующаяся живая система, способная чувствовать, мыслить, активно целенаправленно передвигаться, адаптироваться в среде обитания или приспосабливать ее для удовлетворения своих биологических и социальных потребностей.
Биологической средой обитания называют совокупность природных условий, необходимых для нормального существования живых тел.
Различают внешнюю и внутреннюю биологические среды. Внешняя среда – это комплекс природных факторов, находящихся вне организма, но необходимых для поддержания его жизнедеятельности.
Совокупность биологических жидкостей, омывающих клетки организма, составляют внутреннюю среду организма . Жидкости внутренней среды организма – кровь, лимфа, межклеточная жидкость и другие, являются внешней средой для его клеток.
Живая клетка является элементарной структурно-функциональной единицей организма. Объединение клеток, обеспечивающих выполнение определенной специфической задачи, привело в процессе эволюции к образованию живых тканей – покровной (эпителиальной ), жировой , костной , соединительной , железистой , мышечной и нервной . Живые ткани составляют органы – сердце, легкие, печень, почки и другие, которые обеспечивают выполнение сложных дифференцированных функций, направленных на поддержание жизнедеятельности организма.
Науку о жизнедеятельности здорового организма во взаимодействии его с внешней средой, изучающую функции клеток, тканей, органов, систем органов и организма в целом, а также механизмы их регуляции, называют физиологией (греч. «physis» – природа, «logos» - наука).
Выделяют три основных задачи физиологии:
1) изучение объективных закономерностей протекания физиологических функций (что происходит?),
2) выяснение механизмов реализации физиологических функций (каким образом?),
3) выявление целевого назначения физиологических функций (зачем?).
Физиологическая функция (лат. «functio» - деятельность) – специфическая форма деятельности организма, которая завершается достижением определенного полезного для него результата, позволяющего приспособиться к условиям окружающей среды.
Так, основной функцией сердца является нагнетание крови в сосудистое русло, а желудочно-кишечного тракта – обеспечение поступления питательных веществ в кровь. Большинство органов и систем выполняют несколько функций. Например, основная функция почек - выделение конечных продуктов обмена веществ, однако они также участвуют в регуляции величины артериального давления, осмотического давления и электролитного баланса биологических жидкостей организма.
Физиологические функции являются проявлением жизнедеятельности организма и характеризуются определенными признаками - параметрами . Одна и та же физиологическая функция может быть охарактеризована несколькими параметрами, среди которых выделяют:
1) интенсивность физиологической функции,
2) экстенсивность,
3) мощность,
4) коэффициент полезного действия (КПД),
5) временные характеристики,
6) биоритмы.
Интенсивность характеризует уровень напряженности физиологической функции. Эти параметры выражаются в абсолютной величине качественных проявлений физиологических функций - физиологических констант , т.е. показателей внутренней среды организма (величина артериального давления, температура тела, концентрация глюкозы в крови и другие).
Экстенсивность показывает за счет взаимодействия каких процессов достигается интенсивный параметр. Так, интенсивный параметр функции терморегуляции - температура тела человека, предопределяется экстенсивностью двух разнонаправленных процессов - теплопродукции и теплоотдачи.
Мощность - это работа, совершенная организмом за единицу времени.
КПД (коэффициент полезного действия) - это отношение энергии, затраченной организмом на выполнение полезной работы ко всей израсходованной при этом энергии.
Временные характеристики включают в себя скорость протекания физиологических процессов и их ускорение . Наконец, биоритмы - это периодические циклически повторяющиеся изменения физиологических функций. По продолжительности периода различают три основных вида биоритмов:
1) ультрадианные, с периодичностью от долей секунды до 20 часов,
2) циркадианные (околосуточные) – от 20 до 28 часов,
3) инфрадианные, к которым относят недельные , месячные , сезонные и годовые биоритмы .
Параметры физиологической функции могут изменяться под влиянием регулирующих механизмов . Под биологической регуляцией понимают такое управляемое изменение физиологической функции, которое направлено на обеспечение определенной деятельности живой системы или ее устойчивости к действию возмущающих факторов.
Несмотря на то, что организм постоянно обменивается с окружающей средой веществом, энергией и информацией, любое живое существо способно поддерживать неизменность и стабильность своих внутренних жизненно важных параметров. Относительное постоянство внутренней среды организма и стабильность его физиологических функций называют гомеостазом.
Гомеостаз характеризуется всей совокупностью различных физиологических констант. Такие константы поддерживаются живой системой около уровня, предопределяющего оптимальный клеточный метаболизм - совокупность процессов преобразования веществ и энергии, обеспечивающих жизнедеятельность организма и его взаимосвязь с внешней средой. Поэтому гомеостаз является абсолютно необходимым условием жизни.
Организм является открытой термодинамической системой, которая постоянно обменивается с окружающей средой веществом, энергией и информацией. Поэтому, физиологические показатели не могут быть абсолютно стабильными. Константы, которые изменяются в границах, обеспечивающих биологический оптимум жизнедеятельности и нормальное протекание метаболизма, называются релаксоконстантами .
По амплитуде изменения релаксоконстанты подразделяются на два вида - жесткие и пластичные . Допустимая амплитуда колебания уровня жестких констант невелика. К ним относятся физико-химические показатели внутренней среды, в частности, осмотическое и онкотическое давление плазмы крови, ее кислотно-щелочной баланс. Пластичные константы характеризуются относительно большой допустимой амплитудой колебаний их величины. К ним относятся такие физиологические показатели как, температура тела, артериальное давление, частота сердечных сокращений и дыхания, объем кровотока и множество других.
Для поддержания гомеостаза организм должен:
1) потреблять и расщеплять пищу до питательных веществ, удовлетворяющих энергетические и пластические потребности клеток, тканей и органов,
2) поглощать кислород для окисления питательных веществ, которые являются источником энергии живой системы,
3) выделять в окружающую среду ненужные и вредные продукты обмена веществ,
4) обладать способностью передвигаться, чтобы захватывать пищу, спасаться от врагов.
Если показатели гомеостаза выходят за пределы нормальных диапазонов, определяющих оптимальное протекание метаболизма, то это приводит к нарушению физиологических функций, развитию болезни и гибели.
Для изучения показателей внутренней среды организма и его физиологических функций используют клинические методы исследования и эксперименты . Клинические исследования проводится на человеке, а эксперименты выполняются на животных.
На ранних этапах развития физиологии особо популярны были хирургические экспериментальные методики: экстирпация – удаление части или всего органа и трансплантация – пересадка изучаемого органа в том же организме на новое место или перенос его в другой организм с последующим наблюдением и регистрацией того, какими последствиями сопровождаются такие вмешательства.
С целью изучения деятельности органов, недоступных непосредственному наблюдению, используют фистульныйметод . Он заключается в оперативном создании сообщения внутреннего органа с внешней средой. Разновидностью этой методики может быть катетеризация – введение специальных синтетических трубок в протоки желез или в кровеносные сосуды.
Для того чтобы установить зависимость функции органа от влияния нервной системы, используют методику денервации . С этой целью перерезают нервные волокна, иннервирующие орган.
С появлением современных технических средств появилась инструментальныеметодики - электрофизиологические , биохимические , рентгенологические и многие другие, дающие возможность регистрировать процессы жизнедеятельности без предварительных хирургических операций, что позволяет изучать физиологические функции не только у животных, но и у человека.
В физиологии различают два вида экспериментов, которые связаны с необходимостью выполнять хирургическое вмешательство: острый (вивисекция ) и хронический .
Острый эксперимент характеризуется:
1) отсутствием необходимости соблюдения стерильности в процессе вивисекции,
2) проведением исследования во время или сразу после операции,
3) эвтаназией - умерщвлением животного во время эксперимента или после его завершения.
Острый эксперимент представляет собой достаточно грубое вторжение исследователя в организм. Это необходимо для первоначального накопления данных о функциях органов, тканей и клеток тела. Метод острого эксперимента является методологической основой аналитическогоподхода к изучению функций. Этот подход характеризуется тем, что объектом исследования являются функции отдельных клеток, тканей и органов без учета их взаимосвязи друг с другом и с окружающей средой.
Хронический эксперимент характеризуется:
1) необходимостью соблюдения стерильности во время подготовительной хирургической операции,
2) проведением исследования только после выздоровления животного,
3) многократным изучением физиологических функций органа или организма в целом в условиях максимально приближенных к естественным.
Метод хронического эксперимента является основой системного подхода к исследованию функций организма. Системный методологический подход характеризуется изучением физиологической функции во взаимосвязи с функциональным состоянием организма, как единого целого и с учетом его взаимодействия с внешней средой.
Основоположником системного подхода является русский физиолог, лауреат Нобелевской премии И.П. Павлов. В начале 20-го века он предложил метод физиологической хирургии , позволяющий изучать физиологические функции в условиях естественного поведения, и впервые выдвинул представления о системном характере деятельности организма .
Совокупность органов, выполняющих единую функцию, составляет анатомические системы . У человека имеется девять анатомических систем:
1) опорно-двигательная , образующая остов тела, обеспечивающая движение его частей относительно друг друга и перемещение организма в пространстве;
2) сердечно-сосудистая , обеспечивающая перемещение крови и лимфы по сосудам;
3) дыхательная , необходимая для доставки к тканям кислорода и выведение из организма углекислого газа;
4) пищеварительная , предназначенная для переработки пищи и транспорта образовавшихся питательных веществ в кровь или лимфу;
5) выделительная , обеспечивающая удаление вредных и ненужных для организма продуктов обмена веществ;
6) эндокринная , продуцирующая биологически активные вещества – гормоны, участвующие в регуляции физиологических функций организма;
7) половая , которая выполняет функцию размножения;
8) сенсорная , участвующая в восприятии сигналов из внешнего мира и внутренней среды организма;
9) нервная , объединяющая и регулирующая деятельность всех органов, обеспечивающая взаимодействие организма с внешней средой.
Временное объединение органов, тканей и клеток, связанных единой задачей, которая направлена на получение определенного, полезного для организма результата жизнедеятельности, называют функциональной системой .
Функциональная система (по П.К. Анохину)- это сложный динамический, саморегулирующийся и самообразующийся комплекс, состоящий из центральных и периферических элементов, которые взаимосодействуют друг другу в процессе достижения полезного для организма приспособительного результата, обеспечивающего нормальный метаболизм.
Функциональная система состоит из пяти основных элементов:
1) полезный приспособительный результат (ППР),
2) рецепторы результата,
3) обратная афферентация,
4) нервный центр,
5) исполнительные механизмы.
Конец работы -
Эта тема принадлежит разделу:
Учебное пособие «ОБЩАЯ ФИЗИОЛОГИЯ»
Высшего профессионального образования... ТВЕРСКАЯ ГОСУДАРСТВЕННАЯ МЕДИЦИНСКАЯ АКАДЕМИЯ... МИНИСТЕРСТВА ЗДРАВООХРАНЕНИЯ И СОЦИАЛЬНОГО РАЗВИТИЯ РОССИЙСКОЙ ФЕДЕРАЦИИ...
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ:
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
0001 Физиологические реакции живого организма
Всякий живой организм и все его клетки обладают раздражимостью, т. е. способностью отвечать на воздействия внешней среды или нарушения их состояния изменением своей структуры, возникновением, усилением или ослаблением своей активной деятельности, что неразрывно связано с качественными и количественными изменениями обмена веществ и энергии. Изменения структуры и функций организма и его клеток в ответ на различные воздействия называют биологическими реакциями, а воздействия, их вызывающие, - раздражителями, или стимулами. Понятие биологической реакции – это все виды ответной деятельности организма, его органов и клеток на различные воздействия. Реакции клеток проявляются в изменении их формы, структуры, их роста и процесса деления, в образовании в них различных химических соединений, преобразовании потенциальной энергии в кинетическую (электрическую, механическую, тепловую, световую), совершении той или иной работы (перемещении в пространстве, выделении тех или иных веществ, осмотической работе по концентрированию в клетке определенных электролитов). Реакции целостного организма чрезвычайно многообразны. В процессе их осуществления изменяется деятельность многих органов и бесчисленного множества клеток, ибо организм всегда реагирует на различные воздействия как целое, как единая сложная система. Поэтому хотя реакции организма и совершаются благодаря деятельности клеток, однако они не могут быть сведены к реакциям отдельных клеток. В этом проявляется общее правило, что закономерности системы не могут быть сведены к закономерностям отдельных образующих систему элементов.
Раздражение Раздражителем живой клетки или организма как целого может оказаться любое изменение внешней среды или внутреннего состояния организма, если оно достаточно велико, возникло достаточно быстро и продолжается достаточно долго. Все бесконечное разнообразие возможных раздражителей клеток и тканей можно разделить на три группы: физические, физико-химические и химические. К числу физических раздражителей принадлежат температурные, механические (удар, укол, давление, перемещение в пространстве, ускорение и др.), электрические, световые, звуковые. Физико-химическими раздражителями
являются изменения осмотического давления, активной реакции среды, электролитного состава коллоидального состояния. К числу химических раздражителей относится множество веществ, имеющих различный состав и свойства, изменяющих обмен веществ или структуру клеток. Химическими раздражителями, способными вызывать физиологические реакции, являются поступающие из внешней среды вещества пищи, лекарственные препараты, яды, а также многие химические соединения, образующиеся в организме, например гормоны, продукты обмена веществ. Раздражителями клеток, вызывающими их деятельность, являются нервные импульсы. Нервные импульсы, поступая по нервным волокнам от нервных окончаний в центральную нервную систему или приходя от нее к периферическим органам - мышцам, железам, вызывают изменения их состояния и деятельности. По своему физиологическому значению все раздражители делят на адекватные и неадекватные. Адекватными называются те раздражители, которые действуют на данную биологическую структуру в естественных условиях, к восприятию которых она специально приспособлена и чувствительность к которым у нее чрезвычайно велика. Для палочек и колбочек сетчатки глаза адекватным раздражителем являются лучи видимой части солнечного спектра, для тактильных рецепторов кожи - давление, для вкусовых сосочков языка - разнообразные химические вещества, для скелетных мышц - нервные импульсы, притекающие к ним по моторным нервам. Неадекватными называются те раздражители, для восприятия которых данная клетка или орган специально не приспособлены. Так, мышца сокращается при воздействии кислоты или щелочи, электрического тока, внезапного растяжения, механического удара, быстрого согревания и т. д. Клетки значительно более чувствительны по отношению к своим адекватным раздражителям, чем к неадекватным. Это является выражением функционального приспособления, выработавшегося в процессе эволюции.
Возбудимость Некоторые клетки и ткани (нервная, мышечная и железистая) специально приспособлены к осуществлению быстрых реакций на раздражение. Такие клетки и ткани называют возбудимыми, а их способность отвечать на раздражение возбуждением называют возбудимостью. Мерой возбудимости служит та минимальная сила раздражителя, которая вызывает возбуждение. Эта
минимальная сила раздражения носит название порога раздражения. Чем больше минимальная сила раздражения, требуемая для вызова реакций, чем выше порог раздражения, тем ниже возбудимость, и, наоборот, чем ниже порог раздражения, тем выше возбудимость. По отношению к разным раздражителям порог раздражения может быть различен. Особенно высока возбудимость рецепторов по отношению к адекватным раздражителям, например, для раздражения обонятельной клетки достаточно, чтобы на нее подействовало всего несколько молекул пахучего вещества.
Возбуждение Для возбудимых клеток характерна специфическая форма реагирования на действие раздражителей: в них возникает волнообразный физиологический процесс - возбуждение представляет собой сложную биологическую реакцию, проявляющуюся в совокупности физических, физико-химических, химических процессов и функциональных изменений. Обязательным признаком возбуждения является изменение электрического состояния поверхностной клеточной мембраны. Клетки при возбуждении переходят от состояния физиологического покоя к состоянию свойственной данной клетке физиологической деятельности: мышечное волокно сокращается, железистая клетка выделяет секрет.. В возбудимой клетке постоянно имеется разность электрических потенциалов между ее цитоплазмой и внешней средой, т. е. по обе стороны поверхностной клеточной мембраны. Последняя является, таким образом, поляризованной - ее внутренняя поверхность заряжена отрицательно по отношению к наружной. Эту разность потенциалов называют мембранным потенциалом. Причиной такой разности потенциалов является неравенство концентрации ионов внутри клетки - в ее цитоплазме и снаружи клетки - в окружающей тканевой жидкости: в цитоплазме содержится больше ионов калия и меньше ионов натрия по сравнению с тканевой жидкостью. В состоянии покоя мембрана клетки мало проницаема для ионов Na°. При возбуждении проницаемость мембраны увеличивается, и она пропускает положительно заряженные ионы натрия внутрь клетки, что приводит к понижению мембранной разности потенциалов (деполяризации мембраны) и даже к появлению разности потенциалов противоположного знака. Изменение электрической разности потенциалов при возбуждении получило название потенциала действия. Электрический же ток, возникающий
при соединении возбужденного участка ткани с невозбужденным, называют током действия. Возбуждение представляет собой как бы взрывной процесс, возникающий в результате изменения проницаемости мембраны под влиянием раздражителя. Это изменение вначале относительно невелико и сопровождается лишь небольшой деполяризацией, небольшим уменьшением мембранного потенциала в том месте, где было приложено раздражение, и не распространяется вдоль возбудимой ткани (это так называемое местное возбуждение). Достигнув критического – порогового - уровня, изменение разности потенциалов лавинообразно нарастает и быстро - в нерве за несколько десятитысячных долей секунды - достигает своего максимума. Восстановление исходной разности потенциалов – реполяризация мембраны - происходит вначале за счет выхода ионов калия из клетки. Затем благодаря особому физиологическому механизму, так называемому натрий-калиевому насосу, восстанавливается неравенство ионных концентраций между цитоплазмой и окружающей клетку средой (ионы калия обратно входят в клетку, а ионы натрия выходят из нее). Этот восстановительный процесс требует некоторой затраты энергии, поставщиком которой являются процессы обмена веществ. Характерной особенностью клетки в момент ее возбуждения - в период максимальной деполяризации мембраны - является ее неспособность отвечать на новое раздражение. Состояние невозбудимости клетки во время ее возбуждения носит название рефрактерности. Возбуждение - волнообразно распространяющийся процесс. Возникнув в одной клетке или в одном ее участке, например в одном участке нервного волокна, возбуждение распространяется, переходит на другие клетки или на другие участки той же клетки. Проведение возбуждения обусловлено тем, что потенциал действия, возникший в одной клетке или в одном ее участке, становится раздражителем, вызывающим возбуждение нужных участков. Возбуждение от одной нервной клетки к другой или от нервного волокна к мышечной или железистой клетке передается химическим путем. В нервном окончании образуются химические соединения - передатчики нервного импульса (ацетилхолин, норадреналин и др.), вызывающие возбуждение в той возбудимой клетке, на которой расположено нервное окончание. Химические передатчики нервного импульса называются медиаторами.
Физиология тесно связана с многими другими науками, базируясь в своих исканиях на данных одних наук, и являясь. в свою очередь, основой для развития других. Физика и химия, биофизика и биохимия, анатомия, гистология и эмбриология, общая биология, генетика и кибернетика - вот далеко не полный перечень дисциплин, знание которых имеет большое значение для изучения и понимания физиологических процессов, происходящих в организме.
Однако, ведь Вы готовитесь стать врачами, готовитесь лечить больных людей. А физиология изучает функции нормального. здорового организма - она поэтому и называется "нормальная физиология". Зачем же врачу знать функции здорового организма, если он в своей деятельности будет сталкиваться только с больными? Может быть, было бы достаточно только "патологической физиологии", которая изучается на 3 курсе? Так думают многие студенты, и в итоге получается, что они не имеют прочных знаний ни по нормальной, ни по патологической физиологии, ни по клиническим дисциплинам.
Я думаю, что Вы и сами хорошо себе представляете, для чего врачу надо знать законы и механизмы деятельности здорового человека. Разве станете Вы ремонтировать телевизор, если не знаете принципов его работы, если не представляете, как должен функционировать исправный прибор? Разве станете Вы собирать машину, если не знаете законов взаимодействия всех ее частей и механизмов? А человек - это биологическая машина необозримой сложности, и ремонт ее - очень трудная, деликатная и высококвалифицированная работа! Понять нарушения функций, происходящие в организме при различных заболеваниях, наметить правильные пути их лечения можно только при хорошем знании физиологических процессов, протекающих в нормальном, здоровом организме.
Значение физиологии не исчерпывается изучением нормальных физиологических параметров организма. Одно это помогло бы врачу только установить, имеются ли у человека отклонения от нормы. Но физиология вооружает врача механизмами исправления нарушенных функций, позволяет проводить так называемое "патогенетическое" лечение, устраняющее причину развития болезни, а не ее симптомы. Современный врач не может назначать лечение по принципу "от головы, от живота, от простуды". В процессе исследования больного на основе знания физиологии он должен найти причину болезни и, опять таки опираясь на физиологию, произвести такое вмешательство, которое, и только которое приводит к обязательному выздоровлению.
Знание физиологических методов исследования функций позволяет врачу выбрать такие, которые в данный момент необходимы, помогает правильно оценить результаты этих исследований, и из большого арсенала лекарственных средств и лечебных методов избрать самый действенный в данной ситуации.
Изучение заболеваний человека способствует пониманию многих нормальных физиологических процессов. И.П. Павлов по этому поводу писал: "Мир патологических явлений представляет собой бесконечный ряд всевозможных, особенных, ... не имеющих места в норме комбинаций физиологических явлений. Это как бы ряд физиологических опытов, делаемых природой и жизнью, это часто такое сочетание явлений, которое долго не пришло бы в голову современным физиологам, и которые иногда даже не могли бы быть нарочно воспроизведены техническими средствами современной физиологии". Клиника ставит перед физиологией новые задачи и вместе с тем является богатым источником новых физиологических актов. Значение физиологии для медицины и медицины для физиологии так велико, что только их тесный союз сможет воздвигнуть сложное здание науки о человеческом организме.
Современная физиология является наукой экспериментальной: в основе физиологических исследований лежит опыт, эксперимент. В эксперименте производится искусственное вмешательство в те или иные процессы в организме с целью выяснения роли, значения и взаимной связи отдельных органов и систем. Так, например, при раздражении электрическим током шейного симпатического нерва происходит сужение сосудов уха на оперированной стороне, а перерезка этого нерва вызывает резкое расширение этих сосудов. Эти опыты дали возможность физиологу Клоду Бернару впервые установить нервную регуляцию просвета кровеносных сосудов.
В арсенале физиологии имеются такие методы, как экстирпация, трансплантация, денервация, метод наложения лигатур и анастамозов, фистульные методики, методы раздражения и перерезок, перфузии и регистрации функций и т.д. Большинство указанных методик требует вскрытия животного или хирургической операции. Их применяют в острых или хронических опытах.
Арсенал методов исследования, который используется в опытах на животных, не может быть использован при изучении человеческого организма. Поэтому сведения о функциях многих органов у человека до последнего времени в значительной мере ограничивались данными, полученными в опытах на животных. В настоящее время положение во многом изменилось. Достижения современной физики, радиотехники, электроники м кибернетики дали в руки физиологам такие приборы, которые позволяют, не вмешиваясь в организм человека, регистрировать работу его органов. Многие из этих приборов Вы увидите во время обучения на кафедре нормальной физиологии.
Общие закономерности регуляции функций .
Предметом физиологии являются прежде функции организма и его частей. Поэтому, переходя к изложению физиологии, мы прежде всего должны остановиться на понятиях ОРГАНИЗМ и ФУНКЦИЯ.
ОРГАНИЗМ - это самостоятельно существующая единица органического мира, представляющая собою саморегулирующуюся систему, которая на различные изменения внешней среды реагирует как единое целое, способная существовать лишь при постоянном взаимодействии с окружающей внешней средой и само- возобновляться в результате такого взаимодействия.
Физиологические ФУНКЦИИ - это проявления жизнедеятельности, имеющие приспособительное значение и направленные на достижение определенного полезного для организма результата. Осуществляя различные функции, организм приспосабливается к внешней среде или же приспосабливает среду к своим потребностям.
Основной функцией живого организма является обмен веществ и энергии (метаболизм) . Этот процесс состоит в совокупности химических и физических изменений, в превращении веществ и энергии, постоянно и непрерывно происходящих в организме и во всех его структурах. Метаболизм является необходимым условием жизни. Именно он отличает живое от неживого. Жизнь возможна лишь до тех пор, пока происходит обмен веществ, который поддерживает существование живой протоплазмы и ее самообновление. Прекращение обмена веществ приводит к разрушению протоплазмы и смерти организма.
С обменом веществ связаны все остальные физиологические функции, будь то рост, развитие, размножение, питание, пищеварение, дыхание, выделение, движение или реакции на изменение внешней среды. Деятельность их направлена прежде всего на сохранение оптимальных условий метаболизма. (Обеспечение нормальной работы т.н. метаболического котла). В то же время основу любой функции составляет определенная совокупность превращений веществ и энергии. Это равным образом относится к функциям отдельной клетки, ткани, органа или организма в целом.
Всякий организм, одноклеточный или многоклеточный, нуждается в определенных условиях существования, предоставляемых ему той средой обитания, (т.н. внешней средой) к которой данный вид живых существ приспособился на протяжении всего пути своего эволюционного развития. Функции организма нормально обеспечиваются только при условии, что внешняя среда дает ему возможность получения пищи, при определенной температуре, барометрическом давлении, интенсивности и спектре света и т.п.
При этом следует иметь в виду, что пределы колебаний внешней среды, переносимые организмом высших животных, значительно шире, чем те, которые необходимы для нормального функционирования большинства его клеток. Причина этого состоит в том, что средой обитания для клеток организма является его внутренняя среда, которая изменяется значительно меньше чем внешняя. Внутренней средой организма являются кровь, лимфа и тканевая жидкость, в которых живут клетки.
Функции клеток организма нормальны лишь при относительном постоянстве
осмотического давления, электролитного состава, определенной концентрации водородных ионов, питательных и энергетических ресурсов. Постоянство химического состава и физико-химических свойств внутренней среды является важной особенностью организмов высших животных. Для обозначения этого постоянства Кеннон предложил термин гомеостаз . Выражением гомеостаза является наличие ряда биологических констант , т.е. устойчивых количественных показателей, характеризующих нормальное состояние организма. В зависимости от их значения бывают жесткие и мягкие (пластичные) константы. Отмечая постоянство состава, физико-химических и биологических свойств внутренней среды, следует подчеркнуть, что оно является не абсолютным, а относительным и динамическим. Это постоянство достигается непрерывной работой ряда органов и систем, в результате которой выравниваются происходящие под влиянием изменений внешней среды и жизнедеятельности организма сдвиги в составе и физико-химических свойствах внутренней среды.
Гомеостаз имеет определенные границы. При пребывании особенно длительном, в условиях, значительно отличающихся от тех, к которым организм приспособлен, гомеостаз нарушается и могут произойти сдвиги, не совместимые с жизнью. Даже небольшие нарушения гомеостаза приводят к патологии, поэтому определение относительно постоянных физиологических констант (рН, АД, ЧД,ЧСС, МОД и др.) имеет большое диагностическое значение. Поэтому работа по сохранению гомеостаза совершается постоянно и постоянно регулируется соответствующими регуляторными системами, о которых мы будем говорить позже.
Роль разных органов и их систем в сохранении гомеостаза различна. О них мы также будем говорить в соответствующих разделах курса.
Как уже было сказано, характерной особенностью всякого живого организма является то, что он представляет собой саморегулирующуюся систему, которая реагирует на различные воздействия как единое целое. Принцип саморегуляции заключается в том, что отклонение любой константы от нормального уровня само по себе является сигналом для исправления этих сдвигов. Саморегуляция достигается взаимодействием всех клеток организма, его тканей и органов. Это взаимодействие органов особенно отчетливо выражено в работе т.н. функциональных систем . Такую систему образуют органы, совместная деятельность которых обеспечивает приспособление к определенным условиям среды, обеспечивающее удовлетворение любой внутренней потребности.
Под внутренними потребностями мы впредь будем пронимать всякое более или менее длительное отклонение той или иной константы его внутренней среды от уровня, обеспечивающего нормальную его жизнедеятельность. Именно биологические потребности являются первым толчком в цепи процессов саморегуляции различных функций организма.
Потребностей у живого организма может быть бесчисленное множество. Однако все они объединяются в большие группы - биологические, социальные, половые, пищевые, оборонительные и т.п. Удовлетворение той или иной потребности и представляет для каждого живого организма определенный полезный результат его приспособительной деятельности, т.е. функции.
Регуляция - это направленное изменение функций различных органов и тканей. Все функции в организме регулируются с помощью двух основных регуляторных механизмов - нервного и гуморального .
Гуморальный механизм основан на том, что в различных клетках и органах в ходе процесса обмена веществ образуются различные по своей природе и физиологическому действию химические вещества. Поступая в тканевую жидкость, а затем в кровь, он разносятся по всему организму, и могут оказывать влияние на клетки ткани. Частным случаем гуморальной регуляции является эндокринная, осуществляемая железами внутренней секреции.
Нервный механизм регуляции заключается в том, что по нервам ко всем клеткам и органам посылаются пусковые или модулирующие команды, изменяющие деятельность их в нужном для организма направлении. Изменения состояния одних клеток и органов через посредство нервной системы рефлекторным путем вызывает изменения функций других органов. Этот механизм регуляции является более совершенным, так как взаимодействие клеток через нервную систему осуществляется значительно быстрее, чем гуморально-химическое, и кроме того, нервные импульсы всегда имеют ввиду определенного адресата.
Таким образом, можно выделить два общих принципа регуляции всех функций в организме:
1. Все функции в организме регулируются с помощью нервной или (и) гуморальной системы.
2. Регуляция функций осуществляется по принципу саморегуляции.
Оба эти принципа наиболее ярко и полно проявляются в деятельности так называемых функциональных систем (ФС), которые постоянно образуются при возникновении в организме какой-то потребности и обеспечивают оптимальное ее удовлетворение.
Функциональная система - это совокупность разнородных органов и тканей, объединенных на функциональной основе и обеспечивающих при взаимодействии качественно новые функции и формы деятельности, с результатом, присущим системе в целом и не присущим ее частям в отдельности. ФС - это динамическая, саморегулирующаяся организация, деятельность всех составных элементов которой способствует получению жизненно важного для организма приспособительного результата.
В состав ФС могут входить самые разные органы и ткани, деятельность которых может привести к восстановлению нарушенного гомеостаза. Функциональные системы организуются не по анатомическому, а по физиологическому признаку. Главным системообразующим фактором является цель, результат будущей деятельности ФС. По мере прохождения курса физиологии мы будем характеризовать различные специальные ФС (поддержания рН, осмотического давления, концентрации питательных веществ АД, и т.д.,. Сейчас же мы должны рассмотреть общую схему строения любой ФС. На практических занятиях Вы сами сможете, подставив в эту общую схему специфические регуляторные и исполнительные механизмы и результаты их деятельности, построить схему любой ФС.
Центральным системообразующим фактором каждой ФС является результат ее действия, определяющий в целом для организма нормальные условия течения метаболических процессов. Из этого следует, что именно результат является своеобразной "визитной карточкой" каждой ФС.
В живом организме можно различить 3 группы полезных приспособительных результатов.
Первую группу составляют внутренние константы организма, гомеостатические показатели, определяющие его нормальную жизнедеятельность: рН, концентрация солей, питательных веществ, газов и т.д.
Вторую группу составляют результаты приспособительной деятельности организма в окружающей среде, направленные на удовлетворение его внутренних биологических потребностей, сохранение вида и рода (целенаправленное поведение, приводящее к утолению жажды, голода и т.п.).
Третья группа - результаты социальной деятельности человека, направленные на удовлетворение его социальных потребностей.
В зависимости от свойств результата, ФС будет более простой или более сложной, но общая ее схема остается одинаковой.. Всякое отклонение гомеостатического параметра от нормы (это отклонение называется биологической потребностью, а ее удовлетворение - полезным результатом) немедленно воспринимается рецепторными аппаратами и посредством нервной или (и) гуморальной обратной связи (афферентации) избирательно мобилизует специальные регуляторные аппараты. Последние через исполнительные приборы снова возвращают полезный приспособительный результат к необходимому уровню. Все эти процессы протекают непрерывно с постоянным информированием центра об успехе достижения полезного приспособительного результата. Одни и те же исполнительные механизмы и периферические органы могут быть мобилизованы для выполнения различных функций организма и входить в состав разных ФС.
Живой организм представляет собой сложнейший механизм, состоящий из тесно пригнанных в своей работе различных ФС, имеющих общие точки соприкосновения и определенную иерархию. Однако в каждом конкретном случае, в каждый конкретный момент времени, всегда имеется доминирующая ФС, которая определяет деятельность организма на данный момент и подчиняет себе деятельность других функциональных систем.
Работу многих ФС организма объединяет кровь, как пункт сосредоточения предконечных результатов многих ФС. Поскольку кровь является внутренней средой организма, поскольку ее константы являются гомеостатическими и в значительной мере жесткими, мы и начинаем изучение предмета физиологии с раздела "Физиология системы крови".
ЛЕКЦИИПО ФИЗИОЛОГИИ ЧЕЛОВЕКА
Физиология как наука. Предмет, задачи, методы, история физиологии
Физиология (физис - природа) - это наука о нормальных процессах жизнедеятельности организма, составляющих его физиологических систем, отдельных органов, тканей, клеток и субклеточных структур, механизмах регуляции этих процессов и влиянии на функции организма естественных факторов внешней среды.
Исходя из этого, в целом предметом физиологии является здоровый организм. Задачи физиологии включены в ее определение. Основным методом физиологии является эксперимент на животных. Выделено 2 основных разновидности экспериментов или опытов:
1.Острый опыт или вивисекция (живосечение). В процесс него производится хирургическое вмешательство, исследуются функции открытого или изолированного органа. После этого не добиваются выживания животного. Продолжительность острого эксперимента от нескольких десятков минут до нескольких часов (пример).
2.Хронический опыт. В процессе хронических опытов производят оперативное вмешательство для получения доступности к органу. Затем добиваются заживления операционных ран и лишь после этого приступают к исследованиям. Продолжительность хронических экспериментов может составлять многие годы (пример).
Иногда выделяют подострый эксперимент (пример).
Вместе с тем, для медицины требуются сведения о механизмах функционирования человеческого организма. Поэтому И.П. Павлов писал:” Экспериментальные данные, можно применять к человеку только с осторожностью, постоянно проверяя фактичность сходства с деятельностью этих органов у человека и животных". Следовательно, без постановки специальных наблюдений и опытов на человеке изучение его физиологии бессмысленно. Поэтому выделяют специальную физиологическую науку - физиологию человека, Физиология человека имеет предмет, задачи, методы и историю. Предметом физиологии человека является здоровый человеческий организм.
Её задачи:
1.Исследование механизмов функционирования клеток, тканей, органов, систем организма человека в целом
2. Изучение механизмов регуляции функций органов и систем организма.
3. Выявление реакций человеческого организма и его систем на изменение внешней и внутренней среды.
Так как физиология в целом экспериментальная наука, основным методом физиологии человека также является эксперимент. Однако эксперименты на человеке коренным образом отличаются от опытов на животных. Во-первых, подавляющее большинство исследований на человеке производится с помощью неинвазивных методов, т.е. без вмешательства в органы и ткани (пример ЭКГ, ЭЭГ, ЭМГ, анализы крови и т.д.). Во-вторых, эксперименты на человеке проводят только тогда, когда они не наносят вреда здоровью и с согласия испытуемого. Иногда острые опыты проводятся на человеке в клинике, когда этого требуют задачи диагностики (пример). Однако следует отметить, что без данных классической физиологии возникновение и развитие физиологии человека было бы невозможно (памятники лягушке и собаке). Еще И.П. Павлов, оценивая роль физиологии для медицины, писал: "Понимаемые в грубом смысле слова физиология и медицина не отделимы, знание физиологии необходимо врачу любой специальности". А также, что "Медицина лишь обогащаясь постоянно изо дня в день, новыми физиологическими фактами, станет, наконец тем, чем она должна быть в идеале, т.е. умением чинить испортившийся механизм человека и быть прикладным знанием физиологии" (примеры из клиники). Другой известнейший русский физиолог проф. В.Я. Данилевский отмечал: "Чем точнее и полнее будут определены признаки нормы для телесной и душевной жизни человека, тем правильнее будет диагноз врача для ее патологических отклонений”.
Физиология, являясь основополагающей биологической наукой, тесно связана с другими фундаментальными и биологическими науками. В частности, без знания законов физики невозможно объяснение биоэлектрических явлений, механизмов свето- и звуковосприятия. Без применения данных химии невозможно описание процессов обмена веществ, пищеварения, дыхания и т.д., Поэтому на границах этих наук с физиологией выделились дочерние науки биофизика и биохимия.
Так как структура и функция неразделимы, причем именно функция определяет формирование структуры, физиология тесно связана с морфологическими науками: цитологией, гистологией, анатомией.
В результате исследования действия различных химических веществ на организм из физиологии выделилась в самостоятельные науки фармакология и токсикология. Накопление данных о нарушениях механизмов функционирования организма при различных заболеваниях послужило основой возникновения патологической физиологии.
Выделяют общую и частную физиологию. Общая физиология изучает основные закономерности жизнедеятельности организма, механизмы таких базисных процессов как обмен веществ и энергии, размножение, процессы возбуждения и т.д. Частная физиология исследует функции конкретных клеток, тканей, органов и физиологических систем. Поэтому в ней выделяются такие разделы, как физиология мышечной ткани, сердца, почек, пищеварения, дыхания и т.д. Кроме того, в физиологии выделяют разделы имеющие специфический предмет исследования или особые подходы в исследовании функций. К ним относятся эволюционная физиология (объяснение), сравнительная физиология, возрастная физиология.
В физиологии имеется целый ряд прикладных разделов. Это, например, физиология сельскохозяйственных животных. В физиологии человека выделяют следующие прикладные разделы:
1.Возрастня физиология. Изучает возрастные особенности функций организма.
2.Физиология труда.
3.Клиническая физиология. Это наука, использующая физиологические методики и подходы для диагностики и анализа патологических отклонений.
4.Авиационная и космическая физиология.
5.Физиология спорта.
Физиология человека теснейшим образом связана с такими клиническими дисциплинами, как терапия, хирургия, акушерство, эндокринология, психиатрия, офтальмология и т.д. Например, эти науки используют для диагностики многочисленные методики разработанные физиологами. Отклонения нормальных параметров организма являются основой выявления патологии.
Некоторые разделы физиологии человека являются базой для психологии. Это физиология центральной нервной системы, высшей нервной деятельности, сенсорных систем, психофизиология.
История физиологии подробно описана в учебнике под ред. Ткаченко
^ МЕХАНИЗМЫ РЕГУЛЯЦИИ ФУНКЦИЙ ОРГАНИЗМА
Гуморальная и нервная регуляция. Рефлекс. Рефлекторная дуга. Основные принципы рефлекторной теории
Все функции организма регулируются с помощью двух систем регуляции: гуморальной и нервной. Филогенетически более древняя гуморальная регуляция это регуляция посредством физиологически активных веществ (ФАВ) циркулирующих в жидкостях организма - крови, лимфе, межклеточной жидкости. Факторами гуморальной регуляции являются:
1.Неорганические метаболиты и ионы. Например, катионы кальция, водорода, углекислый газ.
2.Гормоны желез внутренней секреции. Вырабатываются специализированными инкреторными железами. Это инсулин, тироксин и др..
3.Местные или тканевые гормоны. Это гормоны вырабатываются специальными клетками, называемыми паракринными, транспортируются тканевой жидкостью и действуют только на небольшом расстоянии от секретирующих клеток. К ним относятся такие вещества, как гистамин, серотонин, гормоны желудочно-кишечного тракта и другие.
4.Биологически активные вещества, обеспечивающие креаторные связи между клетками ткани. Это белковые макромолекулы, выделяемые ими. Они регулируют дифференцировку, рост и развитие всех клеток составляющих ткань и обеспечивают функциональное объединение клеток в ткань. Такими белками являются, например кейлоны, которые тормозят синтез ДНК и деление клеток.
Основные особенности гуморальной регуляции:
1.Низкая скорость регулирующего воздействия, связанная с невысокой скоростью токов соответствующих жидкостей организма.
2.Медленое нарастание силы гуморального сигнала и медленное снижение. Это связано с постепенным увеличением концентрации ФАВ и постепенным их разрушением.
3.Отсутствие конкретной ткани или органа-мишени для действия гуморальных факторов. Они действуют на все ткани и органы по ходу тока жидкости, в клетках которых имеются соответствующие рецепторы.
Нервная регуляция - это регуляция функций организма посредством рефлексов, осуществляемых нервной системой.
Понятие о рефлекторном принципе деятельности нервной системы впервые разработано в 17 веке французским естествоиспытателем Рене Декартом. Он предложил гипотетическую схему формирования непроизвольного движения (механистическое представление). Термин "рефлекс" (отражательное действие) ввел в физиологию в 1771 году Унзер. Й. Прохаска в 1800 году разработал схему простейшей рефлекторной дуги. И.М. Сеченов распространил понятие "рефлекс" на любую, в том числе и высшую нервную деятельность (ВНД). При этом он исходил из 2-х положений: 1. всякая деятельность организма сводится в конечном итоге к движению. 2. все движения по своему происхождению рефлексы. И.П. Павлов экспериментально обосновал взгляд на рефлекс, как основной акт любой нервной деятельности. Он же разделил все рефлексы, по механизму образования, на безусловные и условные. Основные черты рефлекторной теории И.П. Павлов сформулировал в работе "Ответ физиолога психологам”. Она включает три основополагающих принципа:
1. Принцип детерминизма. Он гласит "нет действия без причины". Т.е. всякий рефлекторный акт является следствием действия раздражителя на организм.
2. Принцип анализа и синтеза. В мозге постоянно происходит анализ, т.е. различение сигналов, а также синтез, т.е. их взаимодействие и целостное восприятие.
3. Принцип структурности. В нервной системе нет процессов, не имеющих определенной структурной локализации.
Морфологической основой любого рефлекса является рефлекторная дуга или рефлекторный путь. Рефлекторная дуга (РД) это путь прохождения рефлекторной реакции, т.е. нервных сигналов. Рефлекторная дуга соматического (двигательного) рефлекса состоит из следующих основных звеньев:
1. Рецептор, воспринимающий раздражение
2. Афферентное или восходящее или чувствительное нервное волокно
3. Нервный центр в Ц.Н.С.
4.Эфферентное или нисходящее, двигательное нервное волокно
5. Исполнительный орган “эффектор”
В ряде рефлекторных дуг имеется нейрон обратной связи (6), или нейрон обратной афферентации, реагирующий на рефлекторный ответ и контролирующий его.
В соматической рефлекторной дуге можно выделить нейроны, выполняющие определенные функции. В частности, в простейшей моносинаптической рефлекторной имеется всего 2 нейрона: чувствительный и двигательный. В рассматриваемой нами простейшей полисинаптической рефлекторной дуге выделяют: а) чувствительный нейрон, б) вставочный нейрон, в) исполнительный нейрон. Рис. В сложных полисинаптических рефлекторных дугах имеется сотни и тысячи нейронов.
В дуге вегетативного рефлекса имеются следующие звенья:
1. Рецептор
2.Афферентное нервное волокно.
3.Нервный центр (например) для симпатических рефлексов в боковых рогах спинного мозга)
4. Преганглионарное нервное волокно
5. Вегетативный ганглий
6.Постганглионарнсе нервное волокно
7.Исполнительный орган. Рис.
Биологические и функциональные системы
В 50-60-х годах канадский биолог Людвиг Берталанфи, используя математические и кибернетические подходы, разработал основные принципы деятельности биологических систем. Они включают:
1. Целостность, т.е. не сводимость свойств системы к простой сумме свойств ее частей. Т.е. невозможно описать свойства биологической системы через функции ее отдельных элементов (пример).
2. Структурность. Возможность объяснения функций системы через ее структуру (пример).
3. Иерархичность, подчиненность элементов системы друг другу сверху вниз. Т.е. вышележащие компоненты системы управляют нижележащими (пример).
4. Взаимосвязь системы и Среды (пример).
Однако Берталанфи не выявил самого главного - системообразующего фактора. Поэтому основная роль в выявлении системных закономерностей, присущих живым организмам принадлежит академику П.К. Анохину. В физиологии давно существовало понятие физиологических систем. Это комплекс морфологически и функционально объединенных органов, имеющих общие механизмы регуляции и выполняющих однородные функции (пример). Однако П.К. Анохин установил, что в организме имеются и другие системы, Например обеспечивающие поддержание жизненно важных параметров организма. Он назвал их функциональными системами (ФУС). По П.К. Анохину ФУС это совокупность органов и тканей, которые обеспечивают достижение цели в определенном виде жизнедеятельности. Эта цель называется полезным приспособительным результатом (ППР). Им может быть какой- либо параметр внутренней среды, например температура тела, нормальное содержание кислорода в крови и т.д., результат поведения, удовлетворяющий биологическую, например пищевую потребность, результат социальной деятельности человека. Для врача важно понимание ФУС, обеспечивающих гомеостаз.
Именно ППР является тем фактором, который объединяет различные органы и системы организма в единое целое - ФУС. Объединение органов в ФУС происходит не по морфологическому, а по функциональному признаку. Поэтому в ФУС могут входить органы и ткани из самых различных физиологических систем. Причем одни и те же органы могут входить сразу в несколько ФУС. Кроме того, в отличие от физиологических систем, ФУС могут быть как наследуемыми, так и формироваться в процессе индивидуальной жизни. Общая схема ФУС для поддержания параметров гомеостаза включает следующие элементы:
2.Рецепторы ППР
3.Афферентный путь
4.Нервный центр
5.Вегетативная регуляция
6.Гуморальная регуляция
7. Поведенческая регуляция
8.Метаболизм (рис.)
Если под влиянием каких-либо причин ППР выходит за границы физиологической нормы, возбуждаются рецепторы ППР. Нервные импульсы от них поступают в нервный центр, регулирующий данную функцию. От него они идут к исполнительным органам, обеспечивающим поддержание соответствующего параметра гомеостаза. Одновременно запускаются гуморальные механизмы регуляции. Когда несмотря на это ППР не приходит к исходному уровню, нервные импульсы от нервного центра начинают поступать в кору больших полушарий. В результате возбуждения ее нейронов включается внешнее звено саморегуляции организма, т.е. поведенческая регуляция. Это целенаправленное изменение поведения живого существа. В результате этих регулирующих воздействий ППР приходит к исходному уровню, т.е. физиологической норме. На ППР непосредственно влияет метаболизм. С другой стороны и сам ППР оказывает прямое воздействие на метаболические процессы. Примеры функционирования различных ФУС.
^ Принципы саморегуляции организма. Понятие о гомеостазе
и гомеокинезе
Способность к саморегуляции - это основное свойство живых систем Оно необходимо для создания оптимальных условий взаимодействия всех элементов, составляющих организм, обеспечения его целостности. Выделяют четыре основных принципа саморегуляции:
1. Принцип неравновесности или градиента. Биологическая сущность жизни заключается в способности живых организмов поддерживать динамическое неравновесное состояние, относительно окружающей среды. Например, температура тела теплокровных выше или ниже окружающей среды. В клетке больше катионов калия, а вне ее натрия и т.д. Поддержание необходимого уровня асимметрии относительно среды обеспечивают процессы регуляции.
2.Принцип замкнутости контура регулирования. Каждая живая система не просто отвечает на раздражение, но и оценивает соответствие ответной реакции действующему раздражению. Т.е. чем сильнее раздражение, тем больше ответная реакция и наоборот. Эта саморегуляция осуществляется за счет обратных положительных и отрицательных обратных связей в нервной и гуморальной системах регуляции. Т.е. контур регуляции замкнут в кольцо. Пример такой связи - нейрон обратной афферентации в двигательных рефлекторных дугах.
3.Принцип прогнозирования. Биологические системы способны предвидеть результаты ответных реакций на основе прошлого опыта. Пример - избегание болевых раздражений после предыдущих.
4. Принцип целостности. Для нормального функционирования живой системы требуется ее структурная целостность.
Учение о гомеостазе было разработано К. Бернаром. В 1878 г. он сформулировал гипотезу об относительном постоянстве внутренней среды живых организмов. В 1929 г. В. Кэннон показал, что способность организма к поддержанию гомеостаза является следствием систем регуляции в организме. Он же предложил термин “гомеостаз”. Постоянство внутренней среды организма (крови, лимфы, тканевой жидкости, цитоплазмы) и устойчивость физиологических функций является результатом действия гомеостатических механизмов. При нарушении гомеостаза, например клеточного, происходит перерождение или гибель клеток. Клеточный, тканевой, органный и другие формы гомеостаза регулируются и координируются гуморальной, нервной регуляцией, а также уровнем метаболизма.
Параметры гомеостаза являются динамическими и в определенных пределах изменяются под влиянием факторов внешней среды (например, рН крови, содержание дыхательных газов и глюкозы в ней и т.д.). Это связано с тем, что живые системы не просто уравновешивают внешние воздействия, а активно противодействуют им. Способность поддерживать постоянство внутренней среды при изменениях внешней, главное свойство отличающее живые организмы от неживой природы. Поэтому они весьма независимы от внешней среды. Чем выше организация живого существа, тем более оно независимо внешней среды (пример).
Комплекс процессов, которые обеспечивают гомеостаз, называется гомеокинезом. Он осуществляется всеми тканями, органами и системами организма. Однако наибольшее значение имеют функциональные системы.
^ Возрастные особенности физиологических функций
В процессе развития организма происходят как количественные, так и качественные изменения. Например, увеличивается количество многих клеток и их размеры. Одновременно, в результате усложнения структуры организма, появляются новые функции. Например, развивающийся мозг ребенка приобретает способность к абстрактному мышлению.
В основе возрастных изменений функций систем организма лежат следующие явления:
1. Неравномерное или гетерохронное созревание органов и систем организма. 2. Этапные возрастные скачки.
3. Акселерация. Т.е. ускорение темпов биологического развития в определенный период.
Созревание отдельных органов и систем происходит не одновременно (гетерохронно). У новорождённого в первую очередь развиваются те физиологические и функциональные системы, которые обеспечивают выживание организма в период перехода от внутриутробного к внеутробному существованию. На основе наблюдений за формированием функциональных систем в процессе онтогенеза, академик П.К. Анохин создал учение о системогенезе. Гетерохронность развития органов и систем можно проиллюстрировать на примере двигательного аппарата ребенка. Первоначально формируются рефлексы обеспечивающие держание головы, затем способность сидеть, затем стоять и, наконец ходить. Программа индивидуального развития выполняется за счет генетического аппарата. На определенных возрастных этапах происходит экспрессия, т.е. активация строго определенных генов. В результате ускоряется созревание той или иной системы, функции организма. Это проявляется возрастным скачком или критическим периодом. Например, скачкообразное изменение структуры и функции органов, систем наблюдается в период полового созревания.
Акселерация связана с воздействием среды и социальных факторов на организм. Она сопровождается 6ыстрым ростом скелета, мышц, внутренних органов, половым созреванием.
Формирование и развитие организма заканчивается примерно к 20 годам. Люди возрасте от 20 до 55-60 лет относят к зрелому возрасту, В этот период все функции организма полностью сформированы, функциональная активность органов и систем находится примерно на одном уровне. Для людей пожилого возраста 65-75 лет характерно возникновение инволюционных перестроек. Одним из основных признаков старения является снижение основного обмена, в результате чего нарушаются метаболические процессы в клетках. Основной обмен уменьшается в результате снижения количества митохондрий в клетках. Считают, что величина основного обмена один из важнейших факторов определяющих продолжительность жизни человека. После 75 лет наступает старость. Резко уменьшается активность всех физиологических процессов. В результате возникают многие старческие болезни, например атеросклероз.
Механизмы нейрогуморальной регуляции с возрастом также изменяются. У новорожденного имеется ограниченное количество сложных безусловных рефлексов, и нет условных. Одновременно клетки высоко чувствительны к гуморальным факторам. По мере роста совершенствуется рефлекторная деятельность ЦНС. В частности уже к году формируются очень сложные рефлексы, обеспечивающие речь. Одновременно снижается первоначальная чувствительность клеток к гуморальным факторам, У зрелого человека имеются высокоорганизованные механизмы нейрогуморальной регуляции. В старческом возрасте скорость и выраженность рефлекторных реакций снижается. Ослабление нервных влияний на органы и ткани обусловлено деструктивными изменениями нервных окончаний и синапсов в ЦНС и на периферии. Одновременно, из-за изменений рецепторного аппарата клеток уменьшается их восприимчивость к ряду гуморальных факторов.
Для педиатрического факультета важно знание периодов детского возраста. Выделяют (по Аршавскому):
1. Период новорожденности 7- 8 дней.
2. Период грудного вскармливания - 5-6 мес.
3. Период смешанного питания с 6 по 12 мес.
4. Период ясельного возраста 1 год - 3 года
5. Период дошкольного возраста 3 - 7 лет.
6. Период младшего школьного возраста 7-12 лет
7. Период старшего школьного возраста 12-17 лет
8, Период юношеского возраста 17-20 лет
Ф И З И О Л О Г И Я И Б И О Ф И З И К А В О З Б У Д И М Ы Х
К Л Е Т О К
^ Понятие о раздражимости, возбудимости и возбуждении. Классификация раздражителей
Раздражимость - это способность клеток, тканей, организма в целом переходить под воздействием факторов внешней или внутренней среды из состояния физиологического покоя в состояние активности. Состояние активности проявляется изменением физиологических параметров клетки, ткани, организма, например изменением метаболизма.
Возбудимость - это способность живой ткани отвечать на раздражение активной специфической реакцией - возбуждением, т.е. генерацией нервного импульса, сокращением, секрецией. Т.е. возбудимость характеризует специализированные ткани - нервную, мышечные, железистые, которые называются возбудимыми. Возбуждение - это комплекс процессов реагирования возбудимой ткани на действие раздражителя, проявляющийся изменением мембранного потенциала, метаболизма и т.д. Возбудимые ткани обладают проводимостью. Это способность ткани проводить возбуждение. Наибольшей проводимостью обладают нервы и скелетные мышцы.
Раздражитель - это фактор внешней или внутренней среды действующий на живую ткань.
Процесс воздействия раздражителя на клетку, ткань, организм называется раздражением.
Все раздражители делятся на следующие группы:
1.По природе
А) физические (электричество, свет, звук, механические воздействия и т.д.)
Б) химические (кислоты, щелочи, гормоны и т.д.)
В) физико-химические (осмотическое давление, парциальное давление газов и т.д.)
Г) биологические (пища для животного, особь другого пола)
Д) социальные (слово для человека).
2.По месту воздействия:
А) внешние (экзогенные)
Б) внутренние (эндогенные)
3.По силе:
А) подпороговые (не вызывающие ответной реакции)
Б) пороговые (раздражители минимальной силы, при которой возникает возбуждение)
В) сверхпороговые (силой выше пороговой)
4.По физиологическому характеру:
А) адекватные (физиологичные для данной клетки или рецептора, которые приспособились к нему в процессе эволюции, например, свет для фоторецепторов глаза).
Б) неадекватные
Если реакция на раздражитель является рефлекторной, то выделяют также:
А) безусловно-рефлекторные раздражители
Б) условно-рефлекторные
^ Законы раздражения. Параметры возбудимости.
Реакция клеток, тканей на раздражитель определяется законами раздражения
1.Закон "все или ничего": При допороговых раздражениях клетки, ткани ответной реакции не возникает. При пороговой силе раздражителя развивается максимальная ответная реакция, поэтому увеличение силы раздражения выше пороговой не сопровождается ее усилением. В соответствии с этим законом реагирует на раздражения одиночное нервное и мышечное волокно, сердечная мышца.
2. 2. Закон силы: Чем больше сила раздражителя, тем сильнее ответная реакция. Однако выраженность ответной реакции растет лишь до определенного максимума. Закону силы подчиняется целостная скелетная, гладкая мышца, так как они состоят из многочисленных мышечных клеток, имеющих различную возбудимость.
3.Закон силы-длительности. Между силой и длительностью действия раздражителя имеется определенная взаимосвязь. Чем сильнее раздражитель, тем меньшее время требуется для возникновения ответной реакции. Зависимость между пороговой силой и необходимой длительностью раздражения отражается кривой силы-длительности. По этой кривой можно определить ряд параметров возбудимости.А) Порог раздражения - это минимальная сила раздражителя, при которой возникает возбуждение.
Б) Реобаза - это минимальная сила раздражителя, вызывающая возбуждение при его действии в течение неограниченно долгого времени. На практике порог и реобаза имеют одинаковый смысл. Чем ниже порог раздражения или меньше реобаза, тем выше возбудимость ткани.
В) Полезное время - это минимальное время действия раздражителя силой в одну реобазу за которое возникает возбуждение.
Г) Хронаксия - это минимальное время действия раздражителя силой в две реобазы, необходимое для возникновения возбуждения. Этот параметр предложил рассчитывать Л. Лапик, для более точного определения показателя времени на кривой силы-длительности. Чем короче полезное время или хронаксия, тем выше возбудимость и наоборот.
В клинической практике реобазу и хронаксию определяют с помощью метода хронаксиметрии для исследования возбудимости нервных стволов.
4. Закон градиента или аккомодации. Реакция ткани на раздражение зависит от его градиента, Т.е. чем быстрее нарастает сила раздражителя во времени тем быстрее возникает ответная реакция. При низкой скорости нарастания силы раздражителя растет порог раздражения. Поэтому если сила раздражителя возрастает очень медленно возбуждения не будет. Это явление называется аккомодацией.
Физиологическая лабильность (подвижность) - это большая или меньшая частота реакций, которыми может отвечать ткань на ритмическое раздражение. Чем быстрее восстанавливается ее возбудимость после очередного раздражения, тем выше ее лабильность. Определение лабильности предложено Н.Е. Введенским. Наибольшая лабильность у нервов, наименьшая у сердечной мышцы.
^ Действие постоянного тока на возбудимые ткани
Впервые закономерности действия постоянного тока на нерв нервно-мышечного препарата исследовал в 19 веке Пфлюгер. Он установил, что при замыкании цепи постоянного тока, под отрицательным электродом, т.е. катодом возбудимость повышается, а под положительным - анодом снижается. Это называется законом действия постоянного тока. Изменение возбудимости ткани (например нерва) под действием постоянного тока в области анода или катода называется физиологическим электротоном. В настоящее время установлено, что под действием отрицательного электрода - катода потенциал мембраны клеток снижается. Это явление называется физическим катэлектротоном. Под положительным - анодом, он возрастает. Возникает физический анэлектртон. Так как, под катодом мембранный потенциал приближается к критическому уровню деполяризации, возбудимость клеток и тканей повышается. Под анодом мембранный потенциал возрастает и удаляется от критического уровня деполяризации, поэтому возбудимость клетки, ткани падает. Следует отметить, что при очень кратковременном действии постоянного тока (1 мсек и менее)МП не успевает измениться, поэтому не изменяется и возбудимость ткани под электродами.
Постоянный ток широко используется в клинике для лечения и диагностики. Например, с помощью него производится электростимуляция нервов и мышц, физиопроцедуры: ионофорез и гальванизация.
^ Строение и функции цитоплазматической мембраны клеток.
Цитоплазматическая клеточная мембрана состоит из трех слоев: наружного белкового, среднего бимолекулярного слоя липидов и внутреннего белкового. Толщина мембраны 7,5-10 нМ. Бимолекулярный слой липидов является матриксом мембраны. Липидные молекулы его обоих слоев взаимодействуют с белковыми молекулами, погруженными в них. От 60 до 75% липидов мембраны составляют фосфолипиды, 15-30% холестерин. Белки представлены в основном гликопротеинами. Различают интегральные белки, пронизывающие всю мембрану и периферические, находящиеся на наружной или внутренней поверхности. Интегральные белки образуют ионные каналы, обеспечивающие обмен определенных ионов между вне- и внутриклеточной жидкостью. Они, также являются ферментами, осуществляющими противоградиентный перенос ионов через мембрану. Периферическими белками являются хеморецепторы наружной поверхности мембраны, которые могут взаимодействовать с различными ФАВ.
Функции мембраны:
Обеспечивает целостность клетки, как структурной единицы ткани.
Осуществляет обмен ионов между цитоплазмой и внеклеточной жидкостью.
4. Производит восприятие и переработку информации поступающей к клетке в виде химических и электрических сигналов.
^ Механизмы возбудимости клеток. Ионные каналы мембраны.
Механизмы возникновения мембранного потенциала (МП) и потенциалов действия (ПД)
В основном, передаваемая в организме информация имеет вид электрических сигналов (например нервные импульсы). Впервые наличие животного электричества установил физиолог Л Гальвани в 1786 г. С целью исследования атмосферного электричества он подвешивал нервно-мышечные препараты лапок лягушек на медном крючке. Когда эти лапки касались железных перил балкона, происходило сокращение мышц. Это свидетельствовало о действии какого-то электричества на нерв нервно-мышечного препарата. Гальвани посчитал, что это обусловлено наличием электричества в самих живых тканях. Однако, А. Вольта установил, что источником электричества является место контакта двух разнородных металлов - меди и железа. В физиологии первым классическим опытом Гальвани считается прикосновение к нерву нервно-мышечного препарата биметаллическим пинцетом, сделанным из меди и железа. Чтобы доказать свою правоту, Гальвани произвел второй опыт. Он набрасывал конец нерва, иннервирующего нервно-мышечный препарат, на разрез его мышцы. В результате возникало ее сокращение. Однако и этот опыт не убедил современников Гальвани. Поэтому другой итальянец Маттеучи произвел следующий эксперимент. Он накладывал нерв одного нервно-мышечного препарата лягушки на мышцу второго, которая сокращалась под действием раздражающего тока. В результате первый препарат тоже начинал сокращаться. Это свидетельствовало о передаче электричества (ПД) от одной мышце к другой. Наличие разности потенциалов между поврежденным и неповрежденным участками мышцы впервые точно установил в 19 веке с помощью струнного гальванометра (амперметра) Маттеучи. Причем разрез имел отрицательный заряд, а поверхность мышцы положительный.
^ Классификация и структура ионных каналов цитоплазматической
мембраны. Механизмы возникновения мембранного потенциала
и потенциалов действия.
Первый шаг в изучении причин возбудимости клеток сделал в своей работе "Теория мембранного равновесия" в 1924 г. английский физиолог Донанн. Он теоретически установил, что разность потенциалов внутри клетки и вне ее, т.е. потенциала покоя или МП, близка к калиевому равновесному потенциалу. Это потенциал, образующийся на полупроницаемой мембране разделяющий растворы с разной концентрацией ионов калия, один из которых содержит крупные непроникающие анионы. Его расчеты уточнил Нернст. Он вывел уравнение диффузионного потенциала. Для калия он будет равен:
Ек=58 lg -------- = 58 lg ----- = - 75 мВ,
Такова теоретически рассчитанная величина мП.
Экспериментально механизмы возникновения разности потенциалов между внеклеточной жидкостью и цитоплазмой, а также возбуждения клеток установили в 1939 году в Кембридже Ходжкин и Хаксли. Они исследовали гигантское нервное волокно (аксон) кальмара и обнаружили, что внутриклеточная жидкость нейрона содержит 400 мМ калия, 50 мМ натрия, 100 мМ хлора и очень мало кальция. Во внеклеточной жидкости содержалось всего 10 мМ калия, 440 мМ натрия, 560 мМ хлора и 10 мМ кальция. Таким образом, внутри клеток имеется избыток калия, а вне их натрия и кальция. Это обусловлено тем, что в клеточную мембрану встроены ионные каналы, регулирующие проницаемость мембраны для ионов натрия, калия, кальция и хлора.
Все ионные каналы подразделяются на следующие группы:
1.По избирательности:
А)Селективные, т.е. специфические. Эти каналы проницаемы для строго определенных ионов.
Б).Малоселективные, неспецифические, не имеющие определенной ионной избирательности. Их в мембране небольшое количество.
2.По характеру пропускаемых ионов:
А) калиевые
Б) натриевые
В) кальциевые
Г) хлорные
3.По скорости инактивации, т.е. закрывания:
А) быстроинактивирующиеся, т.е. быстро переходящие в закрытое состояние. Они обеспечивают быстро нарастающее снижение МП и такое же быстрое восстановление.
Б) медленноинактирующиеся. Их открывание вызывает медленное снижение МП и медленное его восстановление.
По механизмам открывания:
Б) хемозависимые, открывающиеся при воздействии на хеморецепторы мембраны клетки физиологически активных веществ (нейромедиаторов, гормонов и т. д).
В настоящее время установлено, что ионные каналы имеют следующее строение:
1.Селективный фильтр, расположенный в устье канала. Он обеспечивает прохождение через канал строго определенных ионов.
2.Активационные ворота, которые открываются при определенном уровне мембранного потенциала или действии соответствующего ФАВ. Активационные ворота потенциалзависимых каналов имеется сенсор, который открывает их на определенном уровне МП.
3.Инактивационные ворота, обеспечивающие закрывание канала и прекращение проведения ионов по каналу на определенном уровне МП.(Рис).
Неспецифические ионные каналы не имеют ворот.
Селективные ионные каналы могут находиться в трех состояниях, которые определяются положением активационных (м) и инактивационных (h) ворот (рис):
1.Закрытом, когда активационные закрыты, а инактивационные открыты.
2.Активированном, и те и другие ворота открыты.
3.Инактивированном, активационные ворота открыты, а инактивационные закрыты.
Суммарная проводимость для того или иного иона определяется числом одновременно открытых соответствующих каналов. В состоянии покоя открыты только калиевые каналы, обеспечивающие поддержание определенного мембранного потенциала и закрыты натриевые. Поэтому мембрана избирательно проницаема для калия и очень мало для ионов натрия и кальция, за счет имеющихся неспецифических каналов. Соотношение проницаемости мембраны для калия и натрия в состоянии покоя составляет 1:0,04. Ионы калия поступают в цитоплазму и накапливаются в ней. Когда их количество достигает определенного предела, они по градиенту концентрации начинают выходить через открытые калиевые каналы из клетки. Однако уйти от наружной поверхности клеточной мембраны они не могут. Там их удерживает электрическое поле отрицательно заряженных анионов, находящихся на внутренней поверхности. Это сульфат, фосфат и нитрат анионы, анионные группы аминокислот, для которых мембрана не проницаема. Поэтому на наружной поверхности мембраны скапливаются положительно заряженные катионы калия, а на внутренней отрицательно заряженные анионы. Возникает трансмембранная разность потенциалов. Рис.
Выход ионов калия из клетки происходит до тех пор, пока возникший потенциал с положительным знаком снаружи не уравновесит концентрационный градиент калия, направленный из клетки. Т.е. накопившиеся на наружной стороне мембраны ионы калия не будут отталкивать внутрь такие же ионы. Возникает определенный потенциал мембраны, уровень которого определяется проводимостью мембраны для ионов калия и натрия в состоянии покоя. В среднем, величина потенциала покоя близка к калиевому равновесному потенциалу Нернста. Например, МП нервных клеток составляет 55-70 мВ, поперечно-полосатых - 90-100 мВ, гладких мышц - 40-60 мВ, железистых клеток - 20-45 мВ. Меньшая реальная величина МП клеток, объясняется тем, что его величину уменьшают ионы натрия, для которых мембрана незначительно проницаема и они могут входить в цитоплазму. С другой стороны, отрицательные ионы хлора, поступающие в клетку, несколько увеличивают МП.
Так как мембрана в состоянии покоя незначительно проницаема для ионов натрия, необходим механизм выведения этих ионов из клетки. Это связано с тем, что постепенное накопление натрия в клетке привело бы к нейтрализации мембранного потенциала и исчезновению возбудимости. Этот механизм называется натрий-калиевым насосом. Он обеспечивает поддержание разности концентраций калия и натрия по обе стороны мембраны. Натрий-калиевый насос - это фермент натрий-калиевая АТФ-аза. Его белковые молекулы встроены в мембрану. Он расщепляет АТФ и использует высвобождающуюся энергию для противоградиентного выведения натрия из клетки и закачивания калия в неё. За один цикл каждая молекула натрий-калиевой АТФ-азы выводит 3 иона натрия и вносит 2 иона калия. Так как в клетку поступает меньше положительно заряженных ионов, чем выводится из неё, натрий-калиевая АТФ-аза на 5-10 мВ увеличивает мембранный потенциал.
В мембране имеются следующие механизмы трансмембранного транспорта ионов и других веществ:
1.Активный транспорт. Он осуществляется с помощью энергии АТФ. К этой группе транспортных систем относятся натрий-калиевый насос, кальциевый насос, хлорный насос.
2.Пассивный транспорт. Передвижение ионов осуществляется по градиенту концентрации без затрат энергии. Например, вход калия в клетку и выход из неё по калиевым каналам.
3.Сопряженный транспорт. Противоградиентный перенос ионов без затрат энергии. Например таким образом происходит натрий-натриевый, натрий-кальциевый, калий -калиевый обмен ионов. Он происходит за счет разности концентрации других ионов.
Мембранный потенциал регистрируется с помощью микроэлектродного метода. Для этого через мембрану, в цитоплазму клетки вводится тонкий, диаметром менее 1 мкМ стеклянный микроэлектрод. Он заполняется солевым раствором. Второй электрод помещается в жидкость, омывающую клетки. От электродов сигнал поступает на усилитель биопотенциалов, а от него на осциллограф и самописец (рис).
Дальнейшие исследования Ходжкина и Хаксли показали, что при возбуждении аксона кальмара возникает быстрое колебание мембранного потенциала, которое на экране осциллографа имело форму пика (spike). Они назвали это колебание потенциалом действия (ПД). Так как электрический ток для возбудимых мембран является адекватным раздражителем, ПД можно вызвать, поместив на наружную поверхность мембраны отрицательный электрод - катод, а внутреннюю положительный- анод. Это приведет к снижению величины заряда мембраны - ее деполяризации. При действии слабого допорогового тока происходит пассивная деполяризация, т.е. возникает катэлектротон (рис). Если силу тока увеличить до определенного предела, то в конце периода его воздействия на плато катэлектротона появится небольшой самопроизвольный подъём - местный или локальный ответ. Он является следствием открывания небольшой части натриевых каналов, находящихся под катодом. При токе пороговой силы МП снижается до критического уровня деполяризации (КУД), при котором начинается генерация потенциала действия. Он находится для нейронов примерно на уровне - 50 мВ.
На кривой потенциала действия выделяют следующие фазы:
1.Локальный ответ (местная деполяризация), предшествующий развитию ПД.
2.Фаза деполяризации. Во время этой фазы МП быстро уменьшается и достигает нулевого уровня. Уровень деполяризации растет выше 0. Поэтому мембрана приобретает противоположный заряд - внутри она становится положительной, а снаружи отрицательной. Явление смены заряда мембраны называется реверсией мембранного потенциала. Продолжительность этой фазы у нервных и мышечных клеток 1-2 мсек.
3.Фаза реполяризации. Она начинается при достижении определенного уровня МП (примерно +20 мВ). Мембранный потенциал начинает быстро возвращаться к потенциалу покоя. Длительность фазы 3-5 мсек.
4.Фаза следовой деполяризации или следового отрицательного потенциала. Период, когда возвращение МП к потенциалу покоя временно задерживается. Он длится 15-30 мсек.
5.Фаза следовой гиперполяризации или следового положительного потенциала. В эту фазу, МП на некоторое время становится выше исходного уровня ПП. Ее длительность 250-300 мсек.
Амплитуда потенциала действия скелетных мышц в среднем 120-130 мВ, нейронов 80-90 мВ, гладкомышечных клеток 40-50 мВ. При возбуждении нейронов ПД возникает в начальном сегменте аксона - аксонном холмике.
Возникновение ПД обусловлено изменением ионной проницаемости мембраны при возбуждении. В период локального ответа открываются медленные натриевые каналы, а быстрые остаются закрытыми, возникает временная самопроизвольная деполяризация. Когда МП достигает критического уровня, закрытые активационные ворота натриевых каналов открываются и ионы натрия лавинообразно устремляются в клетку, вызывая нарастающую деполяризацию. В эту фазу открываются и быстрые и медленные натриевые каналы. Т.е. натриевая проницаемость мембраны резко возрастает. Причем от чувствительности активационных зависит величина критического уровня деполяризации, чем она выше, тем ниже КУД и наоборот.
Когда величина деполяризация приближается к равновесному потенциалу для ионов натрия (+20 мВ). сила концентрационного градиента натрия значительно уменьшается. Одновременно начинается процесс инактивации быстрых натриевых каналов и снижения натриевой проводимости мембраны. Деполяризация прекращается. Резко усиливается выход ионов калия, т.е. калиевый выходящий ток. В некоторых клетках это происходит из-за активации специальных каналов калиевого выходящего тока. Этот ток, направленный из клетки, служит для быстрого смещения МП к уровню потенциала покоя. Т.е. начинается фаза реполяризации. Возрастание МП приводит к закрыванию и активационных ворот натриевых каналов, что еще больше снижает натриевую проницаемость мембраны и ускоряет реполяризацию.
Возникновение фазы следовой деполяризации объясняется тем, что небольшая часть медленных натриевых каналов остается открытой.
Следовая гиперполяризация связана с повышенной, после ПД, калиевой проводимостью мембраны и тем, что более активно работает натрий-калиевый насос, выносящий вошедшие в клетку во время ПД ионы натрия.
Изменяя проводимость быстрых натриевых и калиевых каналов можно влиять на генерацию ПД, а следовательно на возбуждение клеток. При полной блокаде натриевых каналов, например ядом рыбы тетродонта - тетродотоксином, клетка становится невозбудимой. Это используется в клинике. Такие местные анестетики, как новокаин, дикаин, лидокаин тормозят переход натриевых каналов нервных волокон в открытое состояние. Поэтому проведение нервных импульсов по чувствительным нервам прекращается, наступает обезболивание (анестезия) органа. При блокаде калиевых каналов затрудняется выход ионов калия из цитоплазмы на наружную поверхность мембраны, т.е. восстановление МП. Поэтому удлиняется фаза реполяризации. Этот эффект блокаторов калиевых каналов также используется в клинической практике. Например, один из них хинидин, удлиняя фазу реполяризации кардиомиоцитов, урежает сердечные сокращения и нормализует сердечный ритм.
Также следует отметить, что чем выше скорость распространения ПД по мембране клетки, ткани, тем выше ее проводимость.
^ Соотношение фаз потенциала действия и возбудимости
Уровень возбудимости клетки зависит от фазы ПД. В фазу локального ответа возбудимость возрастает. Это фазу возбудимости называют латентным дополнением.
В фазу реполяризации ПД, когда открываются все натриевые каналы и ионы натрия лавинообразно устремляются в клетку, никакой даже сверхсильный раздражитель не может стимулировать этот процесс. Поэтому фазе деполяризации соответствует фаза полной невозбудимости или абсолютной рефрактерности.
В фазе реполяризации все большая часть натриевых каналов закрывается. Однако они могут вновь открываться при действии сверхпорогового раздражителя. Т.е. возбудимость начинает вновь повышаться. Этому соответствует фаза относительной невозбудимости или относительной рефрактерности.
Во время следовой деполяризации МП находится у критического уровня, поэтому даже допороговые стимулы могут вызвать возбуждение клетки. Следовательно в этот момент ее возбудимость повышена. Эта фаза называется фазой экзальтации или супернормальной возбудимости.
В момент следовой гиперполяризации МП выше исходного уровня, т.е. дальше КУД и ее возбудимость снижена. Она находится в фазе субнормальной возбудимости. Рис. Следует отметить, что явление аккомодации также связано с изменением проводимости ионных каналов. Если деполяризующий ток нарастает медленно, то это приводит к частичной инактивации натриевых, и активации калиевых каналов. Поэтому развития ПД не происходит.
^ ФИЗИОЛОГИЯ МЫШЦ
В организме имеются 3 типа мышц: скелетные или поперечно-полосатые, гладкие и сердечная. Скелетные мышцы обеспечивают перемещение тела в пространстве, поддержание позы тела за счет тонуса мышц конечностей и тела. Гладкие мышцы необходимы для перистальтики органов желудочно-кишечного тракта, мочевыводящей системы, регуляции тонуса сосудов, бронхов и т.д. Сердечная мышца служит для сокращения сердца и перекачивания крови. Все мышцы обладают возбудимостью, проводимостью и сократимостью, а сердечная и многие гладкие мышцы автоматией - способностью к самопроизвольным сокращениям.
^ Ультраструктура скелетного мышечного волокна.
Двигательные единицы Основным морфо-функциональным элементом нервно-мышечного аппарата скелетных мышц является двигательная единица. Она включает мотонейрон спинного мозга с иннервируемыми его аксоном мышечными волокнами. Внутри мышцы этот аксон образует несколько концевых веточек. Каждая такая веточка образует контакт - нервно-мышечный синапс на отдельном мышечном волокне. Нервные импульсы, идущие от мотонейрона, вызывают сокращения определенной группы мышечных волокон.
Скелетные мышцы состоят из мышечных пучков, образованных большим количеством мышечных волокон. Каждое волокно -это клетка цилиндрической формы диаметром 10-100 мкм и длиной от 5 до 400 мкм. Оно имеет клеточную мембрану - сарколемму. В саркоплазме находится несколько ядер, митохондрии, образования саркоплазматического ретикулума (СР) и сократительные элементы - миофибриллы. Саркоплазматический ретикулум имеет своеобразное строение. Он состоит из системы поперечных, продольных трубочек и цистерн. Поперечные трубочки это впячивания саркоплазмы внутрь клетки. К ним примыкают продольные трубочки с цистернами. Благодаря этому, потенциал действия может распространятся от сарколеммы на систему саркоплазматического ретикулума. В мышечном волокне содержится более 1000 миофибрилл, расположенных вдоль него. Каждая миофибрилла состоит из 2500 протофибрилл или миофиламентов. Это нити сократительных белков актина и миозина. Миозиновые протофибриллы толстые, актиновые тонкие.
На миозиновых нитях расположены отходящие под углом поперечные отростки с головками. У скелетного мышечного волокна при световой микроскопии видна поперечная исчерченность, т.е. чередование светлых и темных полос. Темные полосы называют А-дисками или анизотропными, светлые I-дисками (изотропными). В А-дисках сосредоточены нити миозина, обладающие анизотропией и поэтому имеющие темный цвет. I-диски образованы нитями актина. В центре I-дисков видна тонкая Z-пластинка. К ней прикрепляются актиновые протофибриллы. Участок миофибриллы между двумя Z-пластинками называется саркомером. Это структурный элемент миофибрилл. В покое толстые миозиновые нити лишь на небольшое расстояние входят в промежутки между актиновыми. Поэтому в средней части А-диска имеется более светлая Н-зона, где нет актиновых нитей. При электронной микроскопии в ее центре видна очень тонкая М-линия. Она образована цепями опорных белков, к которым крепятся миозиновые протофибриллы (рис).
^ Механизмы мышечного сокращения
При световой микроскопии было замечено, что в момент сокращения ширина А-диска не уменьшается, а I-диски и Н-зоны саркомеров суживаются. С помощью электронной микроскопии установлено, что длина нитей актина и миозина в момент сокращения не изменяется. Поэтому Хаксли и Хэнсон разработали теорию скольжения нитей. Согласно ей мышца укорачивается в результате движения тонких актиновых нитей в промежутки между миозиновыми. Это приводит к укорочению каждого саркомера, образующего миофибриллы. Скольжение же нитей обусловлено тем, что при переходе в активное состояние головки отростков миозина связываются с центрами актиновых нитей и вызывают их движение относительно себя (гребковые движения). Но это последний этап всего сократительного механизма. Сокращение начинается с того, что в области концевой пластинки двигательного нерва возникает ПД. Он с большой скоростью распространяется по сарколемме и переходит с неё по системе поперечных трубочек СР, на продольные трубочки и цистерны. Возникает деполяризация мембраны цистерн и из них в саркоплазму высвобождаются ионы кальция. На нитях актина расположены молекулы еще двух белков -тропонина и тропомиозина. При низкой (менее 10 -8 М) концентрации кальция, т.е. в состоянии покоя, тропомиозин блокирует присоединение мостиков миозина к нитям актина. Когда ионы кальция начинают выходить из СР, молекула тропонина изменяет свою форму таким образом, что освобождает активные центры актина от тропомиозина. К этим центрам присоединяются головки миозина и начинается скольжение за счет ритмического прикрепления и разъединения поперечных мостиков с нитями актина. При этом головки ритмически продвигаются по нитям актина к Z-мембранам. Для полного сокращения мышцы необходимо 50 таких циклов. Передача сигнала от возбужденной мембраны к миофибриллам называется электромеханическим сопряжением. Когда генерация ПД прекращается и мембранный потенциал возвращается к исходному уровню, начинает работать Са-насос (фермент Са-АТФаза). Ионы кальция вновь закачиваются в цистерны саркоплазматического ретикулума и их концентрация падает ниже 10 -8 М. Молекулы тропонина приобретают исходную форму и тропомиозин вновь начинает блокировать активные центры актина. Головки миозина отсоединяются от них и мышца за счет эластичности приходит в исходное расслабленное состояние.
^ Энергетика мышечного сокращения
Источником энергии для сокращения и расслабления служит АТФ. На головках миозина есть каталитические центры, расщепляющие АТФ до АДФ и неорганического фосфата. Т.е. миозин является одновременно ферментом АТФ-азой. Активность миозина как АТФазы значительно возрастает при его взаимодействии с актином. При каждом цикле взаимодействия актина с головкой миозином расщепляется 1 молекула АТФ. Следовательно, чем больше мостиков переходят в активное состояние, тем больше расщепляется АТФ, тем сильнее сокращение. Для стимуляции АТФ-азной активности миозина требуются ионы кальция, выделяющиеся из СР, которые способствуют освобождению активных центров актина от тропомиозина. Однако запасы АТФ в клетке ограничены. Поэтому для восполнения запасов АТФ происходит его восстановление - ресинтез. Он осуществляется анаэробным и аэробным путем. Процесс анаэробного ресинтеза осуществляется фосфагенной и гликолитической системами. Первая использует для восстановления АТФ запасы креатинфосфата. Он расщепляется на креатин и фосфат, который с помощью ферментов переносится на АДФ (АДФ+Ф=АТФ).Фосфагенная система ресинтеза обеспечивает наибольшую мощность сокращения, но в связи с малым количеством креатинфосфата в клетке, она функционирует лишь 5-6 секунд сокращения. Гликолитическая система использует для ресинтеза АТФ анаэробное расщепление глюкозы (гликогена) до молочной кислоты. Каждая молекула глюкозы обеспечивает восстановление трех молекул АТФ. Энергетические возможности этой системы выше, чем фосфагенной, но и она может служить источником энергии сокращения лишь 0,5 - 2 мин. При этом работа гликолитической системы сопровождается накоплением в мышцах молочной кислоты и снижением содержания кислорода. При продолжительной работе, с усилением кровообращения, ресинтез АТФ начинает осуществляться с помощью окислительного фософрилирования, т.е. аэробным путем. Энергетические возможности окислительной системы значительно больше остальных. Процесс происходит за счет окисления углеводов и жиров. При интенсивной работе в основном окисляются углеводы, при умеренной жиры. Для расслабления также нужна энергия АТФ. После смерти содержание АТФ в клетках быстро снижается и когда становится ниже критического, поперечные мостики миозина не могут отсоединиться от актиновых нитей (до ферментативного аутолиза этих белков). Возникает трупное окоченение. АТФ необходима для расслабления потому, что обеспечивает работу Са-насоса.
^ Биомеханика мышечных сокращений.
Одиночное сокращение, суммация, тетанус.
При нанесении на двигательный нерв или мышцу одиночного порогового или сверхпорогового раздражения, возникает одиночное сокращение. При его графической регистрации, на полученной кривой можно выделить три последовательных периода:
1.Латентный период. Это время от момента нанесения раздражения до начала сокращения. Его длительность около 1-2 мсек. Во время латентного периода генерируется и распространяется ПД, происходит высвобождения кальция из СР, взаимодействие актина с миозином и т.д.
2.Период укорочения. В зависимости от типа мышцы (быстрая или медленная) его продолжительность от 10 до 100 мсек.,
3.Период расслабления. Его длительность несколько больше, чем укорочения. Рис.
В режиме одиночного сокращения мышца способна работать длительное время без утомления, но его сила незначительна. Поэтому в организме такие сокращения встречаются редко, например так могут сокращаться быстрые глазодвигательные мышцы. Чаще одиночные сокращения суммируются.
Суммация это сложение 2-х последовательных сокращений при нанесении на нее 2-х пороговых или сверхпороговых раздражений, интервал между которыми меньше длительности одиночного сокращения, но больше продолжительности рефрактерного периода. Различают 2 вида суммации: полную и неполную суммацию. Неполная суммация возникает в том случае, если повторное раздражение наносится на мышцу, когда он уже начала расслабляться. Полная возникает тогда, когда повторное раздражение действует на мышцу до начала периода расслабления, т.е. в конце периода укорочения.(рис 1,2). Амплитуда сокращения при полной суммации выше, чем неполной. Если интервал между двумя раздражениями еще больше уменьшить. Например, нанести второе в середине периода укорочения, то суммации не будет, потому что мышца находится в состоянии рефрактерности.
Тетанус - это длительное сокращение мышцы, возникающее в результате суммации нескольких одиночных сокращений, развивающихся при нанесении на нее ряда последовательных раздражений. Различают 2 формы тетануса: зубчатый и гладкий. Зубчатый тетанус наблюдается в том случае, если каждое последующее раздражение действует на мышцу, когда она уже начала расслабляться. Т.е. наблюдается неполная суммация (рис). Гладкий тетанус возникает тогда, когда каждое последующее раздражение наносится в конце периода укорочения. Т.е. имеет место полная суммация отдельных сокращений и (рис). Амплитуда гладкого тетануса больше, чем зубчатого. В норме мышцы человека сокращаются в режиме гладкого тетануса. Зубчатый возникает при патологии, например тремор рук при алкогольной интоксикации и болезни Паркинсона.
^ Влияние частоты и силы раздражения на амплитуду сокращения
Если постепенно увеличивать частоту раздражения, то амплитуда тетанического сокращения растет. При определенной частоте она станет максимальной. Эта частота называется оптимальной. Дальнейшее увеличение частоты раздражения сопровождается снижением силы
Тетанического сокращения. Частота, при которой начинается снижение амплитуды сокращения, называется пессимальной. При очень высокой частоте раздражения мышца не сокращается (рис.). Понятие оптимальной и пессимальной частот предложил Н.Е.Введенский. Он установил, что каждое раздражение пороговой или сверхпороговой силы, вызывая сокращение, одновременно изменяет возбудимость мышцы. Поэтому при постепенном увеличении частоты раздражения, действие импульсов все больше сдвигаются к началу периода расслабления, т.е. фазе экзальтации. При оптимальной частоте все импульсы действуют на мышцу в фазе экзальтации, т.е. повышенной возбудимости. Поэтому амплитуда тетануса максимальна. При дальнейшем увеличении частоты раздражения, все большее количество импульсов воздействуют на мышцу, находящуюся в фазе рефрактерности. Амплитуда тетануса уменьшается.
Одиночное мышечное волокно, как и любая возбудимая клетка, реагирует на раздражение по закону "все или ничего". Мышца подчиняется закону силы. При увеличении силы раздражения, амплитуда сокращения ее растет. При определенной (оптимальной) силе амплитуда становится максимальной. Если же и дальше повышать силу раздражения, амплитуда сокращения не увеличивается и даже уменьшается за счет катодической депрессии. Такая сила будет пессимальной. Подобная реакция мышцы объясняется тем, что она состоит из волокон разной возбудимости, поэтому увеличение силы раздражения сопровождается возбуждением все большего их числа. При оптимальной силе все волокна вовлекаются в сокращение. Катодическая депрессия - это снижение возбудимости под действием деполяризующего тока - катода, большой силы или длительности.
^ Режимы сокращения. Сила и работа мышц.
Различают следующие режимы мышечного сокращения:
1.Изотонические сокращения. Длина мышцы уменьшается, а тонус не изменяется. В двигательных функциях организма не участвуют.
2.Изометрическое сокращения. Длина мышцы не изменяется, но тонус возрастает. Лежат в основе статической работы, например при поддержании позы тела.
3.Ауксотонические сокращения. Изменяются и длина и тонус мышцы. С помощью их происходит передвижение тела, другие двигательные акты.
Максимальная сила мышц - это величина максимального напряжения, которое может развить мышца. Она зависит от строения мышцы, ее функционального состояния, исходной длины, пола, возраста, степени тренированности человека.
В зависимости от строения, выделяют мышцы с параллельными волокнами (например портняжная), веретенообразные (двуглавая мышца плеча), перистые (икроножная). У этих типов мышц различная площадь поперечного физиологического сечения. Это сумма площадей поперечного сечения всех мышечных волокон, образующих мышцу. Наибольшая площадь поперечного физиологического сечения, а следовательно сила, у перистых мышц. Наименьшая у мышц с параллельным расположением волокон (рис.).
При умеренном растяжение мышцы сила ее сокращения возрастает, но при перерастяжении уменьшается. При умеренном нагревании она также увеличивается, а охлаждении снижается. Сила мышц снижается при утомлении, нарушениях метаболизма и т.д. .Максимальная сила различных мышечных групп определяется динамометрами, кистевым, становым и т.д.
Для сравнения силы различных мышц определяют их удельную или абсолютную силу. Она равна максимальной, деленной на кв. см. площади поперечного сечения мышцы. Удельная сила икроножной мышцы человека составляет 6,2 кг/см2, трехглавой - 16,8 кг/см2, жевательных - 10 кг/см 2.
Работу мышц делят на динамическую и статическую Динамическая выполняется при перемещении груза. При динамической работе изменяется длина мышцы и ее напряжение. Следовательно мышца работает в ауксотоническом режиме. При статической работе перемещения груза не происходит, т.е. мышца работает в изометрическом режиме. Динамическая работа равна произведению веса груза на высоту его подъема или величину укорочения мышцы (А = Р * h). Работа измеряется в кГ.М, джоулях. Зависимость величины работы от нагрузки подчиняется закону средних нагрузок. При увеличении нагрузки работа мышц первоначально растет. При средних нагрузках она становится максимальной. Если увеличение нагрузки продолжается, то работа снижается (рис.). Такое же влияние на величину работы оказывает ее ритм. Максимальная работа мышцы осуществляется при среднем ритме. Особое значение в расчете величины рабочей нагрузки имеет определение мощности мышцы. Это работа выполняемая в единицу времени (Р = А * Т). Вт
^ Утомление мышц
Утомление - это временное снижение работоспособности мышц в результате работы. Утомление изолированной мышцы можно вызвать ее ритмическим раздражением. В результате этого сила сокращений прогрессирующе уменьшается (рис). Чем выше частота, сила раздражения, величина нагрузки тем быстрее развивается утомление. При утомлении значительно изменяется кривая одиночного сокращения. Увеличивается продолжительность латентного периода, периода укорочения и особенно периода расслабления, но снижается амплитуда (рис.). Чем сильнее утомление мышцы, тем больше продолжительность этих периодов. В некоторых случаях полного расслабления не наступает. Развивается контрактура. Это состояние длительного непроизвольного сокращения мышцы. Работа и утомление мышц исследуются с помощью эргографии.
В прошлом веке, на основании опытов с изолированными мышцами, было предложено 3 теории мышечного утомления.
1.Теория Шиффа: утомление является следствием истощения энергетических запасов в мышце.
2.Теория Пфлюгера: утомление обусловлено накоплением в мышце продуктов обмена.
3.Теория Ферворна: утомление объясняется недостатком кислорода в мышце.
Действительно эти факторы способствуют утомлению в экспериментах на изолированных мышцах. В них нарушается ресинтез АТФ, накапливается молочная и пировиноградная кислоты, недостаточно содержание кислорода. Однако в организме, интенсивно работающие мышцы, получают необходимый кислород, питательные вещества, освобождаются от метаболитов за счет усиления общего и регионального кровообращения. Поэтому были предложены другие теории утомления. В частности, определенную роль в утомлении принадлежит нервно-мышечным синапсам. Утомление в синапсе развивается из-за истощения запасов нейромедиатора. Однако главная роль в утомлении двигательного аппарата принадлежит моторным центрам ЦНС. В прошлом веке И.М.Сеченов установил, что если наступает утомление мышц одной руки, то их работоспособность восстанавливается быстрее при работе другой рукой или ногами. Он считал, что это связано с переключением процессов возбуждения с одних двигательных центров на другие. Отдых с включением других мышечных групп он назвал активным. В настоящее время установлено, что двигательное утомление связано с торможением соответствующих нервных центров, в результате метаболических процессов в нейронах, ухудшением синтеза нейромедиаторов, и угнетением синаптической передачи.
^ Двигательные единицы
Основным морфо-функциональным элементом нервно-мышечного аппарата скелетных мышц является двигательная единица (ДЕ). Она включает мотонейрон спинного мозга с иннервируемыми его аксоном мышечными волокнами. Внутри мышцы этот аксон образует несколько концевых веточек. Каждая такая веточка образует контакт - нервно-мышечный синапс на отдельном мышечном волокне. Нервные импульсы, идущие от мотонейрона, вызывают сокращения определенной группы мышечных волокон. Двигательные единицы мелких мышц, осуществляющих тонкие движения (мышцы глаза, кисти), содержат небольшое количество мышечных волокон. В крупных их в сотни раз больше. Все ДЕ в зависимости от функциональных особенностей делятся на 3 группы:
I. Медленные неутомляемые. Они образованы "красными" мышечными волокнами, в которых меньше миофибрилл. Скорость сокращения и сила этих волокон относительно небольшие, но они мало утомляемы. Поэтому их относят к тоническим. Регуляция сокращений таких волокон осуществляется небольшим количеством мотонейронов, аксоны которых имеют мало концевых веточек. Пример - камбаловидная мышца.
IIВ. Быстрые, легко утомляемые. Мышечные волокна содержат много миофибрилл и называются "белыми". Быстро сокращаются и развивают большую силу, но быстро утомляются. Поэтому их называют фазными. Мотонейроны этих ДЕ самые крупные, имеют толстый аксон с многочисленными концевыми веточками. Они генерируют нервные импульсы большой частоты. Мышцы глаза.
IIA. Быстрые, устойчивые к утомлению. Занимают промежуточное положение.
^ Физиология гладких мышц
Гладкие мышцы имеются в стенках большинства органов пищеварения, сосудов, выводных протоков различных желез, мочевыводящей системы. Они являются непроизвольными и обеспечивают перистальтику органов пищеварения и мочевыводящей системы, поддержание тонуса сосудов. В отличие от скелетных, гладкие мышцы образованы клетками чаще веретенообразной формы и небольших размеров, не имеющими поперечной исчерченности. Последнее связано с тем, что сократительный аппарат не обладает упорядоченным строением. Миофибриллы состоят из тонких нитей актина, которые идут в различных направлениях и прикрепляющихся к разным участкам сарколеммы. Миозиновые протофибриллы расположен рядом с актиновыми. Элементы саркоплазматического ретикулума не образуют систему трубочек. Отдельные мышечные клетки соединяются между собой контактами с низким электрическим сопротивлением - нексусами, что обеспечивает распространение возбуждения по всей гладкомышечной структуре. Возбудимость и проводимость гладких мышц ниже, чем скелетных.
Мембранный потенциал составляет 40-60 мВ, так как мембрана ГМК имеет относительно высокую проницаемость для ионов натрия. Причем у многих гладких мышц МП не постоянен. Он периодически уменьшается и вновь возвращается к исходному уровню. Такие колебания называют медленными волнами (МВ). Когда вершина медленные волны достигает критического уровня деполяризации, на ней начинают генерироваться потенциалы действия, сопровождающиеся сокращениями (рис). МВ и ПД проводятся по гладким мышцам со скоростью всего от 5 до 50 см/сек. Такие гладкие мышцы называют спонтанно активными, т.е. они обладают автоматией. Например за счет такой активности происходит перистальтика кишечника. Водители ритма кишечной перистальтики расположены в начальных отделах соответствующих кишок.
Генерация ПД в ГМК обусловлена входом в них ионов кальция. Механизмы электромеханического сопряжения также отличаются. Сокращение развивается за счет кальция, входящего в клетку во время ПД, Опосредует связь кальция с укорочением миофибрилл важнейший клеточный белок - кальмодулин.
Кривая сокращения также отличается. Латентный период, период укорочения, а особенно расслабления значительно продолжительнее, чем у скелетных мышц. Сокращение длится несколько секунд. Гладким мышцам, в отличие от скелетных свойственно явление пластического тонуса. Это способность длительное время находится в состоянии сокращения без значительных энергозатрат и утомления. Благодаря этому свойству поддерживается форма внутренних органов и тонус сосудов. Кроме того гладкомышечные клетки сами являются рецепторами растяжения. При их натяжении начинают генерироваться ПД, что приводит к сокращению ГМК. Это явление называется миогенным механизмом регуляции сократительной активности.