الوظائف الفسيولوجية لجسم الإنسان. الخصائص الفيزيائية والفسيولوجية للعضلات الهيكلية والقلبية والملساء. الكتاب المدرسي "علم وظائف الأعضاء العامة"
محاضرة 1
مقدمة للدورة
موضوع ومهام علم وظائف الأعضاء
تعمد إنشاء أدوية جديدة. لذلك ، يحتاج الطلاب الذين يدرسون الكيمياء الصيدلانية إلى معرفة الأفكار الحديثة حول آليات التأثيرات الدوائية والسامة على الجسم وأن يكونوا قادرين على تطبيق المعرفة المكتسبة في أنشطة الإنتاج المستقبلية.
المفاهيم الفسيولوجية الأساسية
كل الانضباط العلميلديها جهازها المفاهيمي الخاص. بطبيعة الحال ، فإن علم وظائف الأعضاء ، وهو علم نشأ في القرن التاسع عشر ، له مفاهيمه ومصطلحاته الخاصة. فيما يلي أكثرها شيوعًا. سيتم النظر في المصطلحات والمفاهيم الأخرى التي لها معنى أضيق أثناء تقديم أقسام علم وظائف الأعضاء ذات الصلة في المحاضرات اللاحقة.
المفاهيم الفسيولوجية الأساسية.
دور- نشاط محدد لجهاز أو جهاز. على سبيل المثال ، وظائف الجهاز الهضمي هي المحرك ، الإفراز ، الامتصاص ؛ وظيفة الجهاز التنفسي هي تبادل ثاني أكسيد الكربون والأكسجين ؛ وظيفة الجهاز الدوري هي حركة الدم عبر الأوعية. وظيفة عضلة القلب - الانقباض والاسترخاء ، إلخ.
معالجة- مجموعة من الإجراءات المتسلسلة أو الدول التي تهدف إلى تحقيق نتيجة معينة.
آلية- الطريقة التي يتم بها التحكم في العملية أو الوظيفة. ‚علم وظائف الأعضاء عادة ما يميز متوترو الخلطية(أي من خلال إفراز هرمونات أو مواد كيميائية أخرى بيولوجيًا المواد الفعالة) آليات التنظيم.
ص اللائحة- تغيير شدة واتجاه الوظيفة (العملية) من أجل ضمان النشاط الأمثل للأعضاء والأنظمة.
نظامفي علم وظائف الأعضاء ، هي مجموعة من الأعضاء أو الأنسجة أو التكوينات الهيكلية الأخرى المرتبطة بوظيفة مشتركة. على سبيل المثال ، يوفر نظام القلب والأوعية الدموية ، بمساعدة القلب والأوعية الدموية ، إيصال العناصر الغذائية والمواد التنظيمية والوقائية والأكسجين إلى الأنسجة ، فضلاً عن إزالة المنتجات الأيضية ؛ يوفر نظام التصدير (الإفرازي) للخلية ، بمساعدة الشبكة الإندوبلازمية وجهاز جولجي ، تخليق ونقل الهرمونات والبروتينات الدهنية والأسرار الأخرى من الخلايا.
من سر- منتج معين للنشاط الحيوي لخلية تؤدي وظيفة محددة ويتم إطلاقها في البيئة الداخلية للجسم. تسمى عملية توليد السر وعزله إفراز. حسب التركيب ، ينقسم السر إلى بروتين (مصل) ، مخاطي (مخاطي) ، مختلط ودهن.
التحفيز- عوامل البيئة الخارجية والداخلية أو تغيرها والتي تؤثر على الأعضاء والأنسجة مما يؤدي إلى تغيير في نشاط الأخيرة. هناك محفزات ميكانيكية وكهربائية وكيميائية ودرجة الحرارة والصوت وما إلى ذلك. يمكن أن يكون الحافز عتبة، بمعنى آخر. لها تأثير فعال ضئيل ؛ أقصى، زيادة أخرى في قوتها لا تؤدي إلى زيادة رد الفعل المقابل ؛ تغلبت، يمكن أن يكون لعمله تأثير ضار أو مؤلم أو يؤدي إلى أحاسيس غير كافية.
تهيج- تعرض الأنسجة الحية لمؤثرات خارجية وداخلية.
رد فعل- تغير (تكثيف أو إضعاف) نشاط الكائن الحي أو مكوناته استجابة للتهيج.
رد فعل منعكس- إجراء استجابة (أو عملية) في الجسم (جهاز ، عضو ، نسيج ، خلية) ناتج عن رد فعل.
لا ارادي- ظهور أو تغيير في النشاط الوظيفي للأعضاء أو الأنسجة أو الكائن الحي بأكمله ، ويتم ذلك بمشاركة الجهاز العصبي المركزي استجابة لتهيج النهايات العصبية (المستقبلات).
الاهتياجية- قدرة الخلايا الحية على إدراك التغيرات بيئة خارجيةوالاستجابة لهذه التغييرات باستجابة حماسية. كلما انخفضت قوة عتبة الحافز ، زادت استثارة ، والعكس صحيح.
الإثارة- عملية فسيولوجية نشطة تستجيب بها بعض الخلايا الحية (عصبية ، عضلية ، غدية) للتأثيرات الخارجية.
أنسجة قابلة للإثارة- أقمشة قادرة على الاستجابة لتجاوز التأثيرات الخارجية عتبة الإثارة، للانتقال من حالة الراحة الفسيولوجية إلى حالة الإثارة. من حيث المبدأ ، تتمتع جميع الخلايا الحية بدرجة معينة من الاستثارة ، ولكن في علم وظائف الأعضاء ، من المعتاد الإشارة إلى هذه الأنسجة على أنها عصبية وعضلية وغدية.
فسيولوجيا الخلية
أغشية الخلايا.
محاضرة 2
النقل عبر الغشاء.
غشاء المحتملة.
يسمى الفرق في الجهد الكهربائي (بالفولت أو بالسيارات) بين السائل على جانب واحد من الغشاء والسائل على الجانب الآخر غشاء المحتملة(MP) ويشار إليه Vm. عادة ما يكون حجم المجال المغناطيسي للخلايا الحية من -30 إلى -100 ملي فولت ، ويتم إنشاء كل فرق الجهد هذا في المناطق المجاورة مباشرة لغشاء الخلية على كلا الجانبين. يسمى الانخفاض في قيمة MF نزع الاستقطاب، زيادة - فرط الاستقطاب، استعادة القيمة الأصلية بعد إزالة الاستقطاب - عودة الاستقطاب. توجد إمكانات الغشاء في جميع الخلايا ، ولكن في الأنسجة المثيرة (عصب ، عضلي ، غدي) ، إمكانات الغشاء ، أو كما يطلق عليها أيضًا في هذه الأنسجة ، يستريح غشاء المحتملة، يلعب دورًا رئيسيًا في تنفيذ وظائفهم الفسيولوجية. ترجع إمكانات الغشاء إلى خاصيتين رئيسيتين لجميع الخلايا حقيقية النواة: 1) التوزيع غير المتماثل للأيونات بين السوائل خارج الخلايا وداخلها ، بدعم من عمليات التمثيل الغذائي ؛ 2) النفاذية الانتقائية للقنوات الأيونية لأغشية الخلايا.لفهم كيفية نشوء MF ، تخيل أن وعاءًا معينًا ينقسم إلى جزأين بواسطة غشاء لا يمكن اختراقه إلا لأيونات البوتاسيوم. دع الحجرة الأولى تحتوي على 0.1 م ، والثاني 0.01 م كل محلول. نظرًا لأن تركيز أيونات البوتاسيوم (K +) في الحجرة الأولى أعلى بعشر مرات من الثانية ، ففي اللحظة الأولى لكل 10 K + أيونات تنتشر من الحجرة 1 إلى الثانية ، سيكون هناك أيون واحد منتشر في غير إتجاه. نظرًا لأن الأنيونات الكلوريد (Cl-) لا يمكن أن تمر عبر الغشاء مع كاتيونات البوتاسيوم ، فسوف يتشكل فائض من الأيونات الموجبة الشحنة في الحجرة الثانية ، وعلى العكس من ذلك ، ستظهر فائض من الأيونات في الحجرة 1. نتيجة لذلك ، هناك فرق جهد الغشاء، مما يمنع المزيد من انتشار K + في الحجرة الثانية ، حيث يحتاجون إلى التغلب على جاذبية الأيونات السالبة في اللحظة التي يدخلون فيها الغشاء من الحجرة 1 وتنافر الأيونات المماثلة عند الخروج من الغشاء إلى المقصورة 2. وهكذا ، لكل أيون K + يمر عبر الغشاء في هذه اللحظة ، تعمل قوتان - تدرج تركيز كيميائي (أو فرق جهد كيميائي) ، مما يسهل انتقال أيونات البوتاسيوم من الحجرة الأولى إلى الثانية ، و فرق الجهد الكهربائي ، مما يجبر أيونات K + على التحرك في الاتجاه المعاكس. بعد موازنة هاتين القوتين ، يصبح عدد أيونات K + التي تنتقل من الحجرة 1 إلى الحجرة 2 والعكس بالعكس متساويًا ، التوازن الكهروكيميائي. يسمى فرق جهد الغشاء المقابل لمثل هذه الحالة إمكانات التوازن، في هذه الحالة بالذات ، احتمال توازن أيونات البوتاسيوم ( إك). في نهاية القرن التاسع عشر ، أثبت والتر نيرنست أن جهد التوازن يعتمد على درجة الحرارة المطلقة ، وتكافؤ أيون الانتشار ، وعلى نسبة تركيزات هذا الأيون إلى جوانب مختلفةأغشية:
أين السابق-احتمالية التوازن للأيون X ، ص-ثابت الغاز العام = 1.987 كالوري / (درجة مول) ، تيهي درجة الحرارة المطلقة بالدرجات كلفن ، F- رقم فاراداي = 23060 كالوري / بوصة ، ضهي شحنة الأيونات المنقولة ، [X] 1و [x] 2- تركيز الأيونات في الحجرتين 1 و 2.
إذا انتقلنا من اللوغاريتم الطبيعي إلى اللوغاريتم العشري ، فبالنسبة لدرجة حرارة 18 درجة مئوية وأيون أحادي التكافؤ ، يمكن كتابة معادلة نرنست على النحو التالي:
مثال = 0.058 lg
باستخدام معادلة Nernst ، نحسب احتمال توازن البوتاسيوم لخلية خيالية ، بافتراض أن تركيز البوتاسيوم خارج الخلية هو [K +] n \ u003d 0.01 M ، والتركيز داخل الخلايا هو [K +] v \ u003d 0.1 M:
Ek = 0.058 lg = 0.058 lg = 0.058 (-1) = -0.058 ‚= -58 mV
في هذه الحالة، إكسلبي ، لأن أيونات البوتاسيوم ستترك الخلية الافتراضية ، مما يؤدي إلى شحن الطبقة السيتوبلازمية المجاورة للجانب الداخلي للغشاء سلبًا. نظرًا لوجود أيون منتشر واحد فقط في هذا النظام الافتراضي ، فإن إمكانات توازن البوتاسيوم ستكون مساوية لإمكانات الغشاء ( Ek \ u003d Vm).
الآلية المذكورة أعلاه مسؤولة أيضًا عن تكوين إمكانات الغشاء في الخلايا الحقيقية ، ولكن على عكس النظام المبسط المدروس ، حيث يمكن لأيون واحد فقط أن ينتشر عبر الغشاء "المثالي" ، تسمح أغشية الخلية الحقيقية لجميع الأيونات غير العضوية بالمرور من خلالها واحد أو آخر. ومع ذلك ، فكلما قلت نفاذية الغشاء لأي أيون ، قل تأثيره على المجال المغناطيسي. بالنظر إلى هذا الظرف ، غولدمان في عام 1943. تم اقتراح معادلة لحساب قيمة MF للخلايا الحقيقية ، مع مراعاة التركيزات والنفاذية النسبية من خلال غشاء البلازما لجميع الأيونات المنتشرة:
Vm = 0.058 lg
باستخدام طريقة النظائر المسمى ، حدد ريتشارد كينز في عام 1954 نفاذية خلايا عضلات الضفدع للأيونات الأساسية. اتضح أن نفاذية الصوديوم أقل بحوالي 100 مرة من البوتاسيوم ، ولا يساهم Cl-ion بشكل كبير في تكوين MP في خلايا العضلات. لذلك ، بالنسبة لأغشية الخلايا العضلية ، يمكن كتابة معادلة جولدمان بالشكل المبسط التالي:
Vm = 0.058 lg
Vm = 0.058 lg
أظهرت الدراسات التي أجريت باستخدام أقطاب كهربائية دقيقة يتم إدخالها في الخلايا أن إمكانات الراحة لخلايا عضلات الهيكل العظمي للضفدع تتراوح من -90 إلى -100 مللي فولت. يؤكد هذا الاتفاق الجيد بين البيانات التجريبية والنظرية أن إمكانات الراحة يتم تحديدها من خلال تدفقات انتشار الأيونات غير العضوية. في الوقت نفسه ، في الخلايا الحقيقية ، تكون إمكانات الغشاء قريبة من إمكانات توازن الأيون ، والتي تتميز بأقصى نفاذية للغشاء ، أي إمكانات التوازن لأيون البوتاسيوم.
دور الكالسيوم
محاضرة 3
أساسيات الطاقة الحيوية.
الطاقة الكيميائية ، ملامح تكوينها واستخدامها في النظم الحية.
يتم دراسة الآليات الجزيئية لتحويل الطاقة في الخلايا الحية بواسطة الطاقة الحيوية. تتأكسد المركبات العضوية المختزلة ، التي تعمل كمصدر للطاقة للكائنات الحية ، إلى CO 2 و H 2 O ، وتستخدم الطاقة المنبعثة لتخليق ATP وأداء مهام مختلفة.
قبل الشروع في النظر في عمليات طاقة بيولوجية محددة في الخلية ، دعونا نتذكر أنه وفقًا للقانون الثاني للديناميكا الحرارية ، لا يمكن أن تزداد إنتروبيا أي نظام مغلق إلا بمرور الوقت. الانتروبيا هي فئة ديناميكية حرارية تميز درجة الاضطراب في النظام. لذلك ، الزيادة إنتروبيا،في الواقع ، هناك زيادة في درجة اضطراب النظام ، أي الانتقال من هيكل أكثر ترتيبًا إلى هيكل أقل ترتيبًا وانخفاض في المشاركة طاقة مفيدة(أي الطاقة التي يمكن بها العمل). بعبارة أخرى ، يتبع القانون الثاني للديناميكا الحرارية ذلك أي نظام معزول ، يمكن أن تزيد درجة الاضطراب فقط ، ويمكن أن تنخفض كمية الطاقة (المجانية) المفيدة. في الوقت نفسه ، تتطور الكائنات الحية من بويضة مخصبة إلى الحالة بالغتعقيد هيكلها باستمرار. يبدو أن القانون الثاني للديناميكا الحرارية لم يتحقق في النظم الحية. ومع ذلك ، يجب ألا يغيب عن الأذهان أن هذا القانون يشير إلى نظام مغلق ، وأن الكائن الحي هو نظام مفتوح ، يتميز بتبادل مستمر للمادة والطاقة مع البيئة والحفاظ على مستوى منخفض نسبيًا من الانتروبيا عن طريق زيادة في البيئة. القدرة على استخلاص الطاقة المفيدة منها بيئةهي واحدة من تلك الخصائص الرائعة التي تميز الكائنات الحية عن الطبيعة غير الحية. من خلال استهلاك طعام نباتي أو حيواني ، يقوم الكائن الحي بتحويل الجزيئات عالية التنظيم الموجودة فيه إلى ثاني أكسيد الكربون ، H 2 O ومركبات منخفضة الوزن الجزيئي تحتوي على النيتروجين ، والتي تحتوي على أكثر من ذلك بكثير مستوى عالالانتروبيا ، ويتم استخدام الطاقة الناتجة ، بما في ذلك تطور ونمو الكائن الحي الخاص بها ، مصحوبًا بانخفاض في الانتروبيا. المصدر الأساسي للطاقة على الأرض بالنسبة لمعظم الكائنات الحية هو الشمس ، حيث تتزايد إنتروبياها باستمرار ولا يتم التقاط سوى جزء ضئيل من الطاقة المنبعثة في هذه الحالة بواسطة المحيط الحيوي وتستخدمه النباتات وبعض البكتيريا في تفاعلات التمثيل الضوئي ، و الباقي ضاع بشكل لا رجعة فيه في الكون. حتى الحياة الوفيرة المكتشفة مؤخرًا في أعماق قاع المحيطات في منطقة شقوق المحيط على ما يسمى. يعتمد "المدخنون السود" على الشمس لأنها هوائية ، ولا يؤخذ الأكسجين في الغلاف الجوي والمحيطات إلا من خلال عملية التمثيل الضوئي.
في عملية التطور في جسم الحيوانات ، وبطبيعة الحال ، تم تشكيل العديد من الطرق للحصول على الطاقة ونقلها والاستفادة منها ، وفي الغالبية العظمى منها ، تُستخدم مركبات خاصة لتراكم الطاقة الكيميائية - الفوسفاجين، والتي في هيكلها ما يسمى ب عالية الطاقة أو ماكرورجيكالإتصال. الأدينوزين - 5 "- تراي فوسفات (ATP) هو الممثل الأكثر شيوعًا والأكثر أهمية لهذه المجموعة من المركبات. بالإضافة إلى ATP ، يحتوي عدد من البوليفوسفات العضوي الآخر على روابط عالية الطاقة ، على سبيل المثال ، GTP ، CTP. يستخدم الرمز ~ للإشارة إلى رابطة عالية الطاقة. لذلك يتم اختصار جزيء ATP كـ A-R-F ~ F ~ F(لكن-أدينين ، ص-ريبوز). حتى أن بعض البكتيريا تخزن الطاقة في شكل بوليميتافوسفات غير عضوي ، وهو بوليمر خطي بطول غير محدد.
عند درجة الحموضة = 7.0 ، يكون ATP عبارة عن أنيون ذو شحنة عالية. تحتوي مجموعة ثلاثي الفوسفات على 4 مجموعات OH قادرة على التأين. 3 منهم لديهم قيمة pK منخفضة وينفصلون تمامًا عند الرقم الهيدروجيني = 7 ، المجموعة الرابعة تنفصل بنسبة 75 ٪. يعتبر التركيز العالي للشحنات السالبة عاملاً مهمًا في طبيعة الطاقة العالية لـ ATP. هذه الشحنات قريبة من بعضها البعض وهناك تنافر قوي بينهما. مع الانقسام المائي لمجموعة الفوسفات الطرفية ، تضعف قوى التنافر. المنتجات الناتجة لا تميل إلى لم شمل. الميزة الثانية المهمة التي تحدد القيمة السلبية الكبيرة لـ Δ G "يكمن التحلل المائي لـ ATP في حقيقة أن كلا المنتجين من التحلل الدوراني يتم تثبيتهما عن طريق الاقتران. ومع ذلك ، يكاد لا يوجد ATP في الخلايا في شكل أيونات حرة ، ولكن في كثير من الأحيان في شكل معقد مع Mg أو Mn أيونات.
لا ينبغي الخلط بين مفهوم طاقة رابطة الفوسفات ، التي يستخدمها علماء الكيمياء الحيوية ، مع مفهوم طاقة الرابطة ، والتي تعني في الكيمياء الفيزيائية الطاقة اللازمة لكسر الرابطة بين ذرتين في الجزيء. يُفهم أن طاقة رابطة الفوسفات الكبيرة تعني الفرق بين الطاقة الحرة للمواد الأولية والطاقة الحرة لنواتج التفاعل ، والتي يتم خلالها تحلل المركب الفسفوري لتكوين فوسفات غير عضوي.
نقل مجموعة الفوسفات الطرفية ATPعلى الماء يرافقه الافراج طاقة حرة، تقريبًا ضعف ما هو عليه في التحلل المائي لرابطة الفوسفويستر التقليدية ، على سبيل المثال ، في أحادي فوسفات الأدينوزين. طاقة حرةيميز ذلك الجزء من إجمالي الطاقة الموجودة في المادة ، والذي يمكن من خلاله القيام بالعمل فيه الظروف متساوي الحرارة ومتساوي الضغط، بمعنى آخر. عند درجة حرارة وضغط ثابتين ، وهي ظروف نموذجية للأنظمة البيولوجية. يتغيرون طاقة حرةتقاس كيلو كالوري / مولويشار إليه بـ Δ G "إذا استمرت العملية في ظل الظروف القياسية (рН = 7 ، Т = 25 درجة مئوية ، تؤخذ الركائز ومنتجات التفاعل بنفس التركيز الذي يساوي 1 مول / لتر) والرمز Δ جيللتفاعلات في ظل ظروف محددة للأنظمة البيولوجية الحقيقية. في حالة التحلل المائي ATP ، Δ G "\ u003d -7 كيلو كالوري / مول ، و Δ جي\ u003d -11-13 كيلو كالوري / مول تستخدم الكائنات الحية طاقة ATP لأداء الأعمال الميكانيكية للعضلات ، وإنشاء تدرجات التركيز والإمكانيات الكهربائية عبر الغشاء.
سيتم تحديد الآليات المحددة التي تضمن تدفق هذه العمليات في المحاضرات اللاحقة ، والآن سننظر في المبادئ العامة التي تجعل من الممكن استخدام طاقة ATP لأداء الأعمال الكيميائية. تخيل تركيبًا حيويًا نموذجيًا ، أي عملية الابتنائية (الابتنائية من الارتفاع اليوناني ، عمليات التوليف تلك ، بناء الأجزاء الهيكلية للخلية ، على عكس الهدم) ، حيث يجب أن يتحد اثنان من المونومرات -X و Y- مع بعضهما البعض في تفاعل الجفاف (يسمى أيضًا التكثيف) ، يرافقه إطلاق الماء:
X-OH + Y-H X-Y + H 2 O + Δ G (3 كيلو كالوري / مول)
معظم ردود الفعل هذه إندرجونيك(لا ينبغي الخلط بينه وبين ماص للحرارة) ويرافقه زيادة في الطاقة الحرة من قبل Δ جي، والقوة الدافعة لأي تفاعل كيميائي هي الانخفاض طاقة حرةوتتدفق بشكل عفوي قوييُفضل دائمًا التفاعل ، وبالتالي التفاعل التقويضي العكسي (يسمى التحلل المائي) ، لأنه في هذه الحالة يتم إطلاق طاقة حرةتساوي القيمة المطلقة لاستهلاكها في تفاعل التكثيف. لذلك ، في الخلية ، تستمر هذه التفاعلات على عدة مراحل ، ونتيجة لذلك يقترن التركيب غير المواتي بقوة قويرد فعل يشمل ATP. يتم تنفيذ آلية الاقتران على النحو التالي. تفاعل الاندماج المذكور أعلاه ويرافقه زيادة في الطاقة الحرة (افترض Δ G "\ u003d 3 كيلو كالوري / مول) ، مسبوقًا بفسفرة إحدى الركائز بواسطة الجزيء ATP.
A-R-F ~ F ~ F + HON → A-R-F ~ F + X-O-F - G = 4 كيلو كالوري / مول
Δ G "رد الفعل هذا أقل من ذلك بكثير 4 كيلو كالوري / مول) من تفاعلات التحلل المائي ATPونقل مجموعة الفوسفات إلى الماء (حوالي 11 كيلو كالوري / مول). باقي الطاقة ( 7 كيلو كالوري / مول) في جزيء الركيزة المنشط ، والذي يشارك في التفاعل التخليقي اللاحق. ومع ذلك ، فإن الطاقة الحرة لمواد البدء الآن أعلى من الطاقة الحرة لنواتج التفاعل (7 - 3 = 4 كيلو كالوري / مول) ويستمر التفاعل من اليسار إلى اليمين ، أي في اتجاه الاندماج ، مصحوبًا بانخفاض في الطاقة الحرة:
X-O-F + Y-H → X-Y + F n - Δ G = (4 كيلو كالوري / مول)
وبالتالي ، بسبب الطاقة المخزنة في جزيء ATP رد فعل مائييحل محله اثنان متقارن قويتفاعلات.
هناك طريقة أخرى لاستخدام ATP لأداء عمل كيميائي وهي من خلال التصريف المتسلسل ، أو كما يطلق عليه ، التفاعلات الترادفية. في هذه الحالة رد فعل مائيالعائدات بفضل ما يلي قويتفاعلات. ترجع احتمالية حدوث تفاعلات ترادفية إلى حقيقة أن الزيادة في طاقة حرةلا يستبعد حدوث تفاعل كيميائي ، ولكنه يشير فقط إلى أن قيمة ثابت التوازن لمثل هذا التفاعل أقل من الوحدة ، أي إذا كان هناك رد فعل:
أ + ب ج + د + جي "1 (1)
ومن بعد إلى مكافئ= < 1
وبالتالي ، يتحول توازن التفاعل إلى اليسار ، ويصبح هذا التحول أكثر وضوحًا ، وكلما زادت القيمة المطلقة لـ Δ G ". ومع ذلك ، إذا كان التفاعل 1 متبوعًا بالتفاعل 2
С + X CX- Δ ز "2 (2)
ولهذا التفاعل القيمة Δ G "سلبي بشدة ، ثم يتم استهلاك وسيط التفاعل C ، مما يؤدي إلى تحويل توازن التفاعل 1 إلى اليمين. في حالة كانت الطاقة الحرة الكلية للتفاعلين سالبة ، فسوف تستمر في اتجاه استهلاك الركائز A و B وتكوين المنتج CX.
الآن دعونا نلقي نظرة على مكان وجود آليات الحصول على الطاقة ونقل الفوسفاجينات في الخلية من نموذج "غير مشحون" (على سبيل المثال ، ADP) إلى نموذج "مشحون" (ATP) وكيف تعمل.
دورة حمض الكربوكسيل
في معظم خلايا وأنسجة الكائنات الهوائية ، يدخل حمض البيروفيك الناتج عن تحلل السكر مصفوفة الميتوكوندريا، حيث يتم نزع الكربوكسيل منه ، أي ينفصل عنه جزيء من ثاني أكسيد الكربون ، والباقي ثنائي الكربون مجموعة الأسيتيليربط مع أنزيم أ (شهادة توثيق البرامج) ، تشكيل أسيتيل CoA. يتم إنفاق الطاقة المنبعثة نتيجة لهذا التفاعل على نقل جزيء حامل هيدروجين خاص من صورة مؤكسدة إلى شكل مختزل. يسمى ناقل الهيدروجين نيكوتيناميد الأدينين ثنائي النوكليوتيد ويتم اختصاره NAD (في الاعلى) أو NAD + (شكل مؤكسد) و NADH (NADH) أو NADH + H + (شكل مخفض). وهكذا ، فإن جزيئات NAD + وتلك المشابهة لها في التركيب والوظيفة فلافين الأدينين ثنائي النوكليوتيد، مختصر موضة عابرةمثل الفوسفاجين ، فهي قادرة على تخزين الطاقة ونقلها ، ولكن ليس في شكل رابطة فوسفات عالية الطاقة ، ولكن في شكل ذرات هيدروجين نشطة. أسيتيل- CoA المتكون من حمض البيروفيك يخضع لمزيد من الأكسدة إلى دورة حمض الكربوكسيل. نتيجة التفاعل الأول للدورة (الشكل) من الأسيتيل- C وحمض أوكسالاسيتيك (في الوسط المائي يوجد في شكل أيون - أوكسالأسيتات) يحتوي على 4 ذرات كربون بمشاركة إنزيم سينثيز السترات ، ثلاثي الكربوكسيل ، ستة كربون حمض الليمون(سترات). لذلك ، تسمى دورة حمض الكربوكسيل أحيانًا دورة حمض الستريك.ثم يتم تحويل حامض الستريك إلى العزلة(isocitrate). نتيجة لتفاعل نزع الكربوكسيل التالي ، يتم تحويل جزيء ستة كربون لحمض الأيزوسيتريك إلى خمسة كربون حمض ألفا أوكسوجلوتاريك(α- أوكسوجلوتارات) و NAD + يتم تقليله إلى NADH. علاوة على ذلك ، يتم نزع الكربوكسيل من حمض ألفا أوكسوجلوتاريك إلى العنبر(سكسينات). في الوقت نفسه ، لا يتم تبديد الطاقة المنبعثة على شكل حرارة ، ولكن يتم إنفاقها على اختزال جزيء NAD + آخر إلى NADH وتشكيل منتج وسيط برابطة فوسفات عالية الطاقة ، مما يضمن الفسفرة. غوانوزين ثنائي فوسفات(الناتج المحلي الإجمالي) في غوانوزين ثلاثي الفوسفات(GTP).
نتيجة للتفاعلات الثلاثة التالية ، يتحول حمض السكسينيك على التوالي إلى حمض فوماريك (فومارات), حمض الماليك(مالات) و أوكسو أسيتيكويتم ترميمه بواسطة جزيء FAD و NAD.
من وجهة نظر الطاقة الحيوية ، في كل منعطف من الدورة ، تتم إزالة ذرتين من الكربون دخلت الدورة كـ acetyl-CoA في شكل CO 2 ويتم إنشاء 8 [H] + 2 [H] عند الإدخال - هذا هو مصدر طاقة الهيدروجين المنشط.
وهكذا ، فإن التفاعلات الأنزيمية لدورة حمض الكربوكسيليك تستمر بطريقة تجعل ركيزة التفاعل الأول حمض أوكسالاسيتيكهو أيضًا نتاج التفاعل الثامن الأخير ، أي يتم تجديده بعد كل ثورة في الدورة وتبدأ العملية مرة أخرى. نتيجة للدورة الأولية ، يتم تحويل ذرتين من الكربون الموجودتين في acetyl-CoA إلى CO 2. تم تحديد الطبيعة الدورية لتسلسل تفاعل أكسدة أسيتيل CoA ومراحلها الرئيسية في عام 1937 من قبل عالم الكيمياء الحيوية الإنجليزي من أصل ألماني ، الحائز على جائزة نوبل (1953) هانز كريبس. لذلك ، فإن عملية أكسدة الأسيتيل CoA لها اسم آخر - دورة كريبس. نظرًا لحقيقة أن الأكسدة تحدث تدريجيًا ، من خلال سلسلة من التغييرات الصغيرة نسبيًا في الطاقة الحرة ، فإن الطاقة المنبعثة لا تتبدد على شكل حرارة غير مجدية ، ولكن يتم إنفاقها على اختزال ثلاثة جزيئات NAD إلى NADH وجزيء FAD واحد إلى FADH و تشكيل رابطة فوسفات عالية الطاقة. تتكون هذه الرابطة من فسفرة غوانوزين ثنائي فوسفات إلى غوانوزين ثلاثي الفوسفات. هذا الأخير يتبادل بسهولة مجموعة الفوسفات مع ADP لتشكيل ATP.
يجب أن يؤخذ في الاعتبار أنه ، إلى جانب تحلل السكر ، فإن العمليات الكيميائية الحيوية الأخرى التي تحدث في الخلايا ، على وجه الخصوص ، أكسدة الأحماض الأمينية والأحماض الدهنية (وكذلك الإيثانول) ، هي مصدر أسيتيل CoA للأكسدة في TCA. تدخل أربعة جزيئات NADH ، بما في ذلك الجزيء المتكون أثناء أكسدة البيروفات إلى acetyl-CoA ، وجزيء FADH في سلسلة الجهاز التنفسي ، حيث تتأكسد ذرات الهيدروجين التي تحملها بواسطة الأكسجين الجزيئي إلى الماء. يتم إنفاق الطاقة المنبعثة في هذه الحالة على تخليق ATP من ADP والفوسفات غير العضوي نتيجة لأكثر العمليات تعقيدًا وغموضًا إلى حد كبير ، والتي تسمى الفسفرة التأكسدية.
محاضرة 4
المحاضرة 5
المحاضرة 6
الجسيمات الحالة: الهيكل وطرق التكوين في الخلية ، التصنيف.
الليزوزومات هي مجموعة غير متجانسة (غير متجانسة) من الهياكل الشبيهة بالفجوة السيتوبلازمية بحجم 1-3 ميكرومتر ، وتتمثل السمة المميزة لها في وجود بيئة حمضية وعدد كبير من هيدرولازات مختلفة - إنزيمات قادرة على الانقسام (التحلل المائي) الرئيسي أنواع الجزيئات الكبيرة. يحدد وجود الإنزيمات المائية في الجسيمات الحالة وظيفتها الرئيسية في الخلية - تقسيم الجزيئات الكبيرة والتكوينات الأكبر ، التي تدخل الخلية من الفضاء خارج الخلية وتلك ذات الأصل داخل الخلية. ومن المثير للاهتمام أن النباتات لا تحتوي على الجسيمات الحالة.
تُعرف ثلاثة مسارات محتملة لتكوين الجسيمات الحالة في الخلية. في كل حالة ، يتم تشكيل تشكيلات مختلفة شكليًا ، مما يؤدي إلى فصل المواد عن مصادر مختلفة.
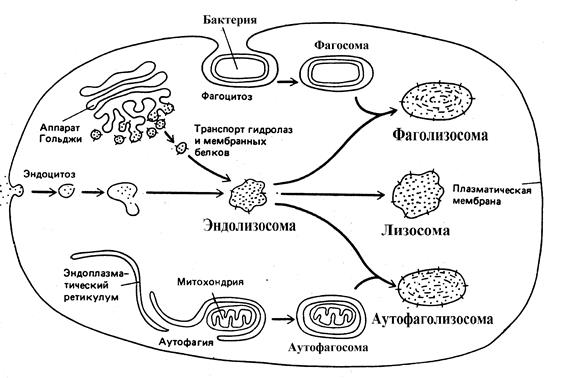 أرز. 6 طرق لتكوين الجسيمات الحالة في الخلية
أرز. 6 طرق لتكوين الجسيمات الحالة في الخلية
في الحالة الأولى ، تدخل المادة القابلة للانقسام - البروتينات أو عديد النيوكليوتيدات أو السكريات - إلى الخلية عن طريق الالتقام. خلال هذه العملية ، تُحاط الجزيئات الكبيرة بما يكفي وغير القادرة على اختراق الأغشية تدريجيًا بجزء صغير من غشاء البلازما ، الذي ينفصل أولاً (ينفصل) ثم ينقسم إلى الخلية ، مكونًا حويصلة تحتوي على المادة التي تلتقطها الخلية . تسمى الفقاعات التي تشكلت نتيجة الالتقام الخلوي اندوسومات. عندما ينتقل الجسيم الداخلي من غشاء الخلية إلى داخل الخلية ، فإنه يتفاعل بشكل متكرر مع حويصلات النقل التي تخرج من نشوة- سطح جهاز جولجي الإنزيمات المتحللة للماء والبروتينات الغشائية ، يتحول إلى الجسيم الداخلي. تستغرق عملية تكوين وتحويل الجسيم الداخلي حوالي 15 دقيقة ويصاحبها تحمض البيئة الداخلية بسبب ضخ أيونات H + من العصارة الخلوية إلى الجسيم الداخلي بواسطة مضخة البروتون المعتمدة على ATP ، والتي تعمل مثل ATP-ase من غشاء الميتوكوندريا الداخلي.
يسمى المسار الثاني لتشكيل الجسيمات الحالة الالتهام الذاتي. في عملية الالتهام الذاتي ، يتم تدمير الأجزاء المستهلكة من الخلية نفسها. من المعروف ، على سبيل المثال ، أن متوسط عمر الميتوكوندريا الواحدة في خلايا الكبد هو حوالي 10 أيام ، وبعد ذلك يجب استخدامها في الجسيمات الحالة. على الصور المجهرية الإلكترونية للخلايا الطبيعية ، يمكن للمرء أن يرى الجسيمات الحالة التي تحتوي على الميتوكوندريا في مراحل مختلفة من التدهور. لقد سبق أن لوحظ أعلاه أنه عن طريق الالتهام الذاتي ، يتم إزالة فائض من ER السلس من خلايا الكبد بعد التوقف عن تناول وإفراز المواد الغريبة الحيوية - المحفزات من الجسم. يبدو أن عملية الالتهام الذاتي تبدأ بمحيط العضية بواسطة الأغشية الموردة من ER ، مما يؤدي إلى تكوين البلعمة الذاتية. بعد ذلك ، يُعتقد أن جسيم البلعمة يندمج مع الجسيم الداخلي ، ويشكل جسيمًا ذاتيًا ، حيث تحدث عملية تدهور جزء ER أو عضية أخرى.
المسار الثالث لتكوين الجسيمات الحالة متاح فقط في الخلايا المتخصصة البلعمةالجسيمات الكبيرة والكائنات الدقيقة. يمكن لمثل هذه الخلايا البلعمية ، والتي تشمل خلايا الدم - العدلات والخلايا الأحادية ، أن تمتص الأجسام الكبيرة من الفضاء خارج الخلية ، وتشكل البلعمة. ثم يصبح البلعوم البلعمةبنفس طريقة البلعمة الذاتية ، أي الانصهار مع الجسيم الداخلي.
غالبًا ما يشار إلى Endosomes و autophagosomes و phagosomes بالمصطلح الشائع - prelysosomes، و endolysosomes ، autophagolysosomes و phagolysosomes تسمى lysosomes. في الجسيمات الحالة الناضجة ، تتحلل المادة الممتصة إلى جزيئات فردية ، مثل الأحماض الأمينية ، التي تدخل العصارة الخلوية وتشارك في التحولات الكيميائية الحيوية اللاحقة. لا تتعرض شظايا الغشاء البلازمي للهيدرولازات ويتم إعادتها مرة أخرى إلى غشاء البلازما بمساعدة حويصلات النقل ، حتى قبل التكوين النهائي للجسيم الحال. تبقى المنتجات غير القابلة للهضم وتتراكم في الجسيمات الحالة التي تفقد الإنزيمات المتحللة للماء وتتحول إلى postlysosomesأو الجثث المتبقية. مع التقدم في السن ، في الخلايا البشرية والحيوانية ، عدد الأجسام المتبقية التي تحتوي على عدد كبير من ليبوفوسينأو صبغة الشيخوخة.
Lipofuscin هو بوليمر حيوي ذو طبيعة مختلفة ، غير قابل لمزيد من الانقسام بسبب روابط كيميائيةبين المونومرات الفردية لم تتشكل في التفاعلات الكيميائية الحيوية العادية ، ولكن نتيجة لعمليات الأكسدة العفوية ، وخاصة الجذور الحرة. تؤدي الأمراض المختلفة والتعرض للإشعاع والعوامل البيئية السلبية الأخرى إلى تسريع تراكم صبغة الشيخوخة.
نواة الخلية والجينوم
المحاضرة 7
محاضرة 1
مقدمة للدورة
موضوع ومهام علم وظائف الأعضاء
علم وظائف الأعضاء (من الكلمات اليونانية: physis - الطبيعة ، الشعارات - التدريس ، العلم) هو علم وظائف كل من الجسم ككل ومكوناته الفردية: الأعضاء والأنسجة والخلايا ؛ العمليات والآليات التي تجعل من الممكن تنفيذ وتنظيم هذه الوظائف وتضمن النشاط الحيوي للإنسان والحيوان في تفاعلهم مع البيئة. تتمثل مهمة علم وظائف الأعضاء في إنشاء الأسس الفيزيائية والكيميائية لعمليات الحياة على جميع مستويات تنظيم الأنظمة الحية من الخلايا الفرعية إلى مستوى الكائن الحي بأكمله. فيما يتعلق بتعقيد موضوع الدراسة ، ينقسم علم وظائف الأعضاء إلى عدة أقسام. وتشمل هذه: علم وظائف الأعضاء العام والخلوي ، وتتمثل مهمتها في دراسة أنماط عمليات الحياة الأساسية والآليات المميزة لجميع الأنظمة الوظيفية والهيكلية للجسم ، وعلم وظائف الأعضاء الخاص ، والذي ينقسم بدوره إلى فسيولوجيا الأعضاء الفردية: القلب والكلى والكبد وما إلى ذلك ؛ فسيولوجيا الأنظمة الوظيفية: التنفس ، والدورة الدموية ، والهضم ، ونظام الغدد الصماء ، وعلم وظائف الأعضاء التنموي ، وعلم وظائف الأعضاء العصبية وفسيولوجيا السلوك ، وعلم وظائف الأعضاء الحسية. هناك أيضًا فسيولوجيا تطبيقية: الرياضة ، والعمل ، والتغذية ، وفسيولوجيا الفضاء وتحت الماء.
ما هي الأهمية العملية لعلم وظائف الأعضاء؟ لماذا من المهم معرفة كيف يعمل الجسم؟ السبب الأكثر وضوحًا هو أن علم وظائف الأعضاء ، وخاصة دراسة أداء جسم الإنسان ، هو الأساس العلمي للطب والكيمياء الصيدلانية. في جميع المراحل السابقة لتطور الحضارة ، كان نهج علاج الأمراض تجريبيًا بالكامل تقريبًا ، أي نزل إلى التجربة والخطأ. نتيجة لذلك ، لوحظ تدريجيًا أن بعض العلاجات تساعد في علاج أمراض معينة - سواء كانت مستحضرات كيميائية أو عشبية ، أو مياه معدنية وحمامات ساخنة ، أو الوخز بالإبر ، أو حتى طرق التأثير النفسي التي كانت شائعة الاستخدام في الطب الشعبي. تأثير الشفاء للكثيرين الأدوية الحديثةتم اكتشافه لأول مرة تجريبيًا بحتًا (على سبيل المثال ، النتروجليسرين ، الذي استخدمه نوبل لإنتاج الديناميت ، مع التسبب في انخفاض حاد في ضغط الدم لدى العمال ، وغالبًا ما يؤدي إلى الوفاة) ، ولا تزال الآليات الرئيسية الكامنة وراء عمل الشفاء لعدد منهم تبقى مجهولة. ومع ذلك ، مع تطور فهمنا لآليات عمل وعلم أمراض الأعضاء والأنسجة وتعميقه ، يتناقص دور النتائج التجريبية البحتة ويصبح من الممكن تطوير طرق فعالة قائمة على الأدلة لعلاج الأمراض التي تصيب الإنسان ، عمدا خلق
1.1. منظر عامعن علم وظائف الأعضاء
رئيسي أسئلة: الكائن الحي والموئل. فهم علم وظائف الأعضاء. الوظيفة الفسيولوجية وخصائصها متعددة العوامل. الاستتباب وثوابت الاسترخاء وأنواعها. طرق البحث في علم وظائف الأعضاء. التجارب الحادة والمزمنة. المناهج التحليلية والنظامية لدراسة الوظائف الفسيولوجية. نظرية النظم الوظيفية وفقًا لـ P.K. أنوخين.
كائن بشري- نظام حي شمولي منظم ذاتيًا قادرًا على الشعور والتفكير والتحرك بنشاط هادف والتكيف مع البيئة أو تكييفها لتلبية احتياجاتها البيولوجية والاجتماعية.
الموائل البيولوجيةدعا مجموع الظروف الطبيعية اللازمة للوجود الطبيعي للأجسام الحية.
هناك بيئات بيولوجية خارجية وداخلية. بيئة خارجيةعبارة عن مجموعة من العوامل الطبيعية التي توجد خارج الجسم ، ولكنها ضرورية للحفاظ على نشاطه الحيوي.
مجموع السوائل البيولوجية التي تستحم خلايا الجسم هي البيئة الداخلية للجسم . سوائل البيئة الداخلية للجسم - الدم والليمفاوية والسوائل بين الخلايا وغيرها ، هي البيئة الخارجية لخلايا الجسم.
خلية حيةهي وحدة هيكلية ووظيفية أولية للجسم. أدى توحيد الخلايا التي تضمن أداء مهمة محددة في عملية التطور إلى التكوين الأنسجة الحية – ساترة (طلائية), دهني, عظم, الضامة, غدي, عضليو متوتر. الأنسجة الحية جثث- القلب والرئتين والكبد والكلى وغيرها ، والتي توفر تنفيذ وظائف متمايزة معقدة تهدف إلى الحفاظ على النشاط الحيوي للجسم.
يُطلق على علم النشاط الحيوي للكائن الحي السليم في تفاعله مع البيئة الخارجية ، والذي يدرس وظائف الخلايا والأنسجة والأعضاء وأنظمة الأعضاء والجسم ككل ، وكذلك آليات تنظيمها. علم وظائف الأعضاء (اليونانية "physis" - الطبيعة ، "الشعارات" - العلم).
هناك ثلاث مهام رئيسية لعلم وظائف الأعضاء:
1) دراسة الأنماط الموضوعية لمسار الوظائف الفسيولوجية (ما الذي يحدث؟) ،
2) توضيح آليات تنفيذ الوظائف الفسيولوجية (كيف؟) ،
3) تحديد الغرض المقصود من الوظائف الفسيولوجية (لماذا؟).
الوظيفة الفسيولوجية(اللاتينية "functio" - النشاط) - شكل معين من نشاط الجسم ، والذي ينتهي بتحقيق نتيجة معينة مفيدة له ، مما يسمح له بالتكيف مع الظروف البيئية.
لذا ، فإن الوظيفة الرئيسية للقلب هي ضخ الدم في قاع الأوعية الدموية ، والجهاز الهضمي هو ضمان إمداد الدم بالعناصر الغذائية. تؤدي معظم الأجهزة والأنظمة وظائف عديدة. على سبيل المثال ، الوظيفة الرئيسية للكلى هي إفراز المنتجات النهائية الأيضية ، ولكنها تشارك أيضًا في تنظيم ضغط الدم ، والضغط الاسموزي ، وتوازن السوائل في الجسم.
الوظائف الفسيولوجية هي مظهر من مظاهر النشاط الحيوي للجسم وتتميز بعلامات معينة - المعلمات. يمكن وصف الوظيفة الفسيولوجية نفسها بعدة معايير ، من بينها:
1) شدة الوظيفة الفسيولوجية ،
2) التمدد ،
3) القوة ،
4) عامل الكفاءة (COP) ،
5) خصائص الوقت ،
6) النظم الحيوية.
الشدةيميز مستوى شدة الوظيفة الفسيولوجية. يتم التعبير عن هذه المعلمات في القيمة المطلقة للمظاهر النوعية للوظائف الفسيولوجية - الثوابت الفسيولوجية، بمعنى آخر. مؤشرات البيئة الداخلية للجسم (ضغط الدم ، درجة حرارة الجسم ، تركيز الجلوكوز في الدم ، وغيرها).
اتساعيظهر بسبب تفاعل العمليات التي يتم تحقيق المعلمة المكثفة. لذلك ، فإن المعلمة المكثفة لوظيفة التنظيم الحراري - درجة حرارة جسم الإنسان ، محددة مسبقًا باتساع عمليتين موجهتين بشكل مختلف - إنتاج الحرارة ونقل الحرارة.
قوةهو العمل الذي يقوم به الكائن الحي لكل وحدة زمنية.
نجاعة(الكفاءة) هي نسبة الطاقة التي ينفقها الجسم لأداء عمل مفيد إلى كل الطاقة التي ينفقها في هذه الحالة.
توقيتتضمن سرعةمسار العمليات الفسيولوجية و التسريع. أخيراً، إيقاعات بيولوجية - هذه تغيرات دورية دورية متكررة في الوظائف الفسيولوجية. وفقًا لمدة الفترة ، هناك ثلاثة أنواع رئيسية من الإيقاع الحيوي:
1) ultradian ، بتردد كسور من الثانية إلى 20 ساعة ،
2) الساعة البيولوجية (على مدار الساعة) - من 20 إلى 28 ساعة ،
3) إنفراديان ، والتي تشمل أسبوعي, فترة, موسميو إيقاعات بيولوجية سنوية.
قد تتغير معلمات الوظيفة الفسيولوجية تحت تأثير الآليات التنظيمية. تحت التنظيم البيولوجي فهم مثل هذا التغيير الخاضع للرقابة في الوظيفة الفسيولوجية ، والتي تهدف إلى ضمان نشاط معين لنظام حي أو مقاومته لعمل العوامل المزعجة.
على الرغم من حقيقة أن الجسم يتبادل باستمرار المادة والطاقة والمعلومات مع البيئة ، فإن أي كائن حي قادر على الحفاظ على ثبات واستقرار معاييره الحيوية الداخلية. يسمى الثبات النسبي للبيئة الداخلية للجسم واستقرار وظائفه الفسيولوجية التوازن.
يتميز الاستتباب بمجمل الثوابت الفسيولوجية المختلفة. يتم الحفاظ على هذه الثوابت من خلال نظام حي بالقرب من المستوى الذي يحدد مسبقًا الخلية المثلى التمثيل الغذائي - مجموعة من عمليات تحويل المواد والطاقة التي تضمن النشاط الحيوي للكائن وعلاقته بالبيئة الخارجية. لذلك ، فإن التوازن هو شرط ضروري للغاية للحياة.
الكائن الحي هو نظام ديناميكي حراري مفتوح يتبادل باستمرار المادة والطاقة والمعلومات مع البيئة. لذلك ، لا يمكن أن تكون المعلمات الفسيولوجية مستقرة تمامًا. تسمى الثوابت التي تتغير ضمن الحدود التي تضمن المستوى البيولوجي الأمثل للحياة والمسار الطبيعي لعملية التمثيل الغذائي ثوابت الاسترخاء .
حسب سعة التغيير تنقسم ثوابت الاسترخاء إلى نوعين - قاسٍو بلاستيك. سعة تذبذب المستوى المسموح به ثوابت صلبة صغير. وتشمل هذه المعلمات الفيزيائية والكيميائية للبيئة الداخلية ، على وجه الخصوص ، الضغط التناضحي والورم في بلازما الدم ، وتوازنها الحمضي القاعدي. الثوابت البلاستيكية تتميز بالسعة الكبيرة المسموح بها نسبيًا للتقلبات في حجمها. وتشمل هذه المؤشرات الفسيولوجية مثل درجة حرارة الجسم وضغط الدم ومعدل ضربات القلب والتنفس وتدفق الدم وغيرها الكثير.
للحفاظ على التوازن ، يجب على الجسم:
1) تستهلك وتحطيم الطعام إلى مغذيات تلبي احتياجات الخلايا والأنسجة والأعضاء من الطاقة والبلاستيك ،
2) امتصاص الأكسجين لأكسدة العناصر الغذائية التي تعد مصدر الطاقة للنظام الحي ،
3) إطلاق منتجات التمثيل الغذائي غير الضرورية والضارة في البيئة ،
4) لديه القدرة على التحرك من أجل التقاط الطعام ، للهروب من الأعداء.
إذا تجاوزت مؤشرات الاستتباب النطاقات الطبيعية التي تحدد المسار الأمثل لعملية التمثيل الغذائي ، فإن هذا يؤدي إلى انتهاك الوظائف الفسيولوجية وتطور المرض والموت.
لدراسة مؤشرات البيئة الداخلية للجسم ووظائفه الفسيولوجية ، طرق البحث السريريو التجارب. يتم إجراء الدراسات السريرية على البشر ، ويتم إجراء التجارب على الحيوانات.
في المراحل الأولى من تطور علم وظائف الأعضاء ، كانت التقنيات الجراحية التجريبية شائعة بشكل خاص: استئصال إزالة جزء أو كل عضو و الزرع - زرع العضو قيد الدراسة في نفس الكائن الحي إلى مكان جديد أو نقله إلى كائن حي آخر ، ثم مراقبة وتسجيل نتائج مثل هذه التدخلات.
من أجل دراسة أنشطة الأعضاء التي يتعذر الوصول إليها للمراقبة المباشرة ، فإنها تستخدم طريقة الناسور . وهو يتألف من الإنشاء السريع لرسالة من الجهاز الداخلي مع البيئة الخارجية. يمكن أن يكون الاختلاف في هذه التقنية قسطرة - إدخال أنابيب تركيبية خاصة في مجاري الغدد أو الأوعية الدموية.
من أجل إثبات اعتماد وظيفة العضو على تأثير الجهاز العصبي ، يتم استخدام تقنية التعصيب . لهذا الغرض ، يتم قطع الألياف العصبية التي تعصب العضو.
مع ظهور التكنولوجيا الحديثة ، طرق مفيدة - الكهربية, البيوكيميائية, إشعاعيوغيرها الكثير ، مما يجعل من الممكن تسجيل العمليات الحيوية دون عمليات جراحية أولية ، مما يجعل من الممكن دراسة الوظائف الفسيولوجية ليس فقط في الحيوانات ، ولكن أيضًا في البشر.
في علم وظائف الأعضاء ، هناك نوعان من التجارب المرتبطة بالحاجة إلى إجراء التدخل الجراحي: حار(تشريح) و مزمن.
تجربة حادةتتميز:
1) عدم الحاجة للمحافظة على العقم في عملية تشريح الأحياء ،
2) إجراء البحوث أثناء الجراحة أو بعدها مباشرة ،
3) القتل الرحيم- قتل الحيوان أثناء التجربة أو بعد إتمامها.
التجربة الحادة هي تدخل فظ إلى حد ما من الباحث في الجسم. هذا ضروري للتراكم الأولي للبيانات حول وظائف أعضاء وأنسجة وخلايا الجسم. طريقة التجربة الحادة هي الأساس المنهجي النهج التحليلي لدراسة الوظائف. يتميز هذا النهج بحقيقة أن موضوع الدراسة هو وظائف الخلايا والأنسجة والأعضاء الفردية دون مراعاة علاقتها مع بعضها البعض ومع البيئة.
تجربة مزمنةتتميز:
1) ضرورة المحافظة على العقم أثناء التحضير عملية جراحية,
2) إجراء البحوث فقط بعد شفاء الحيوان ،
3) دراسة متكررة للوظائف الفسيولوجية لعضو أو كائن حي ككل في ظل ظروف قريبة قدر الإمكان من الطبيعة.
طريقة التجربة المزمنة هي الأساس نهج النظملدراسة وظائف الجسم. يتميز النهج المنهجي المنهجي بدراسة الوظيفة الفسيولوجية فيما يتعلق بالحالة الوظيفية للجسم ككل ، مع مراعاة تفاعله مع البيئة الخارجية.
مؤسس نهج النظام هو عالم وظائف الأعضاء الروسي ، الحائز على جائزة نوبل I.P. بافلوف. في بداية القرن العشرين ، اقترح طريقة الجراحة الفسيولوجية، مما يجعل من الممكن دراسة الوظائف الفسيولوجية في ظروف السلوك الطبيعي ، ولأول مرة طرح أفكار حولها الطبيعة الجهازية لنشاط الجسم.
مجموعة من الأعضاء التي تؤدي وظيفة واحدة هي أنظمة تشريحية . يمتلك الإنسان تسعة أنظمة تشريحية:
1) الجهاز العضلي الهيكلي ، التي تشكل الهيكل العظمي للجسم ، مما يضمن حركة أجزائه بالنسبة لبعضها البعض وحركة الجسم في الفضاء ؛
2) القلب والأوعية الدموية ، توفير حركة الدم والليمفاوية عبر الأوعية ؛
3) تنفسي ضروري لتوصيل الأكسجين إلى الأنسجة وإزالة ثاني أكسيد الكربون من الجسم ؛
4) الجهاز الهضمي مصمم لمعالجة الطعام ونقل العناصر الغذائية الناتجة إلى الدم أو اللمف ؛
5) مطرح الذي يضمن إزالة المنتجات الأيضية الضارة وغير الضرورية للجسم ؛
6) الغدد الصماء ، إنتاج مواد نشطة بيولوجيا - هرمونات تشارك في تنظيم الوظائف الفسيولوجية للجسم ؛
7) جنسي التي تؤدي وظيفة التكاثر ؛
8) حسي تشارك في إدراك الإشارات من العالم الخارجي والبيئة الداخلية للجسم ؛
9) متوتر ، الذي يوحد وينظم نشاط جميع الأجهزة ، مما يضمن تفاعل الكائن الحي مع البيئة الخارجية.
يسمى الارتباط المؤقت للأعضاء والأنسجة والخلايا المتصلة بمهمة واحدة ، والتي تهدف إلى الحصول على نتيجة معينة من النشاط الحيوي المفيد للجسم ، نظام وظيفي .
نظام وظيفي(وفقًا لـ P.K. Anokhin) هو مركب ديناميكي معقد ، منظم ذاتيًا وتشكيلًا ذاتيًا ، يتكون من عناصر مركزية ومحيطية تتفاعل مع بعضها البعض في عملية تحقيق نتيجة تكيفية مفيدة للجسم وتضمن التمثيل الغذائي الطبيعي.
يتكون النظام الوظيفي من خمسة عناصر رئيسية:
1) نتيجة تكيفية مفيدة (PPR) ،
2) مستقبلات النتيجة ،
3) التوكيد العكسي ،
4) مركز العصب ،
5) الآليات التنفيذية.
نهاية العمل -
هذا الموضوع ينتمي إلى:
الكتاب المدرسي "الفسيولوجيا العامة"
أعلى فائق التعليم المهني... الأكاديمية الطبية لولاية TVER ... التابعة لوزارة الصحة والتنمية الاجتماعية للاتحاد الروسي ...
إذا كنت بحاجة إلى مواد إضافية حول هذا الموضوع ، أو لم تجد ما كنت تبحث عنه ، فإننا نوصي باستخدام البحث في قاعدة بيانات الأعمال لدينا:
ماذا سنفعل بالمواد المستلمة:
إذا كانت هذه المادة مفيدة لك ، فيمكنك حفظها في صفحتك على الشبكات الاجتماعية:
0001 التفاعلات الفسيولوجية للكائن الحي
كل كائن حي وكل خلاياه لديه قابلية للتهيج ، أي القدرة على الاستجابة للتأثيرات أو الاضطرابات البيئية في حالته عن طريق تغيير بنيته ، وظهور نشاطه القوي أو تقويته أو إضعافه ، والذي يرتبط ارتباطًا وثيقًا بالتغيرات النوعية والكمية في التمثيل الغذائي والطاقة. التغييرات في بنية ووظائف الجسم وخلاياه استجابة للتأثيرات المختلفة تسمى التفاعلات البيولوجية ، والتأثيرات التي تسببها تسمى المنبهات أو المنبهات. مفهوم التفاعل البيولوجي هو جميع أنواع نشاط استجابة الجسم وأعضائه وخلاياه للتأثيرات المختلفة. تتجلى تفاعلات الخلايا في تغيير شكلها وهيكلها وعملية نموها وانقسامها ، في تكوين مركبات كيميائية مختلفة فيها ، وتحويل الطاقة الكامنة إلى حركية (كهربائية ، ميكانيكية ، حرارية ، ضوئية) ، الأداء عمل واحد أو آخر (التحرك في الفضاء ، إطلاق مواد معينة ، العمل التناضحي على تركيز بعض الإلكتروليتات في الخلية). ردود فعل الكائن الحي متنوعة للغاية. في عملية تنفيذها ، يتغير نشاط العديد من الأعضاء والخلايا التي لا تعد ولا تحصى ، لأن الجسم يتفاعل دائمًا مع التأثيرات المختلفة ككل ، كنظام معقد واحد. لذلك ، على الرغم من أن تفاعلات الكائن الحي تتم بسبب نشاط الخلايا ، إلا أنه لا يمكن اختزالها في تفاعلات الخلايا الفردية. يوضح هذا القاعدة العامة التي تقول إن انتظامات النظام لا يمكن اختزالها في انتظام العناصر الفردية التي تشكل النظام.
تهيجيمكن أن يكون مصدر إزعاج لخلية حية أو كائن حي ككل أي تغيير في البيئة الخارجية أو الحالة الداخليةالكائن الحي ، إذا كان كبيرًا بدرجة كافية ، نشأ بسرعة كافية ويستمر لفترة كافية. يمكن تقسيم كل الأنواع اللانهائية من المهيجات المحتملة للخلايا والأنسجة إلى ثلاث مجموعات: فيزيائية ، وفيزيائية - كيميائية ، وكيميائية. تشمل المنبهات الفيزيائية درجة الحرارة ، والميكانيكية (الصدمة ، والوخز ، والضغط ، والحركة في الفضاء ، والتسارع ، وما إلى ذلك) ، والكهرباء ، والضوء ، والصوت. المحفزات الفيزيائية والكيميائية
هي التغيرات في الضغط الاسموزي ، التفاعل النشط للوسط ، تكوين المنحل بالكهرباء للحالة الغروية. تشمل المهيجات الكيميائية العديد من المواد ذات التركيب والخصائص المختلفة التي تغير التمثيل الغذائي أو بنية الخلايا. المهيجات الكيميائية التي يمكن أن تسبب تفاعلات فسيولوجية هي المواد الغذائية القادمة من البيئة الخارجية والأدوية والسموم وكذلك العديد من المركبات الكيميائية التي تتكون في الجسم ، مثل الهرمونات ومنتجات التمثيل الغذائي. المحفزات الخلوية التي تسبب نشاطها هي نبضات عصبية. النبضات العصبية ، التي تأتي على طول الألياف العصبية من النهايات العصبية للجهاز العصبي المركزي أو القادمة منها إلى الأعضاء المحيطية - العضلات والغدد ، تسبب تغيرات في حالتها ونشاطها. وفقًا لأهميتها الفسيولوجية ، يتم تقسيم جميع المحفزات إلى كافية وغير كافية. كافية هي تلك المحفزات التي تعمل على بنية بيولوجية معينة في ظروف طبيعية ، لإدراكها الذي تم تكييفها بشكل خاص والحساسية التي تكون عالية للغاية تجاهها. بالنسبة لقضبان الشبكية ومخاريطها ، فإن أشعة الجزء المرئي من الطيف الشمسي هي مصدر تهيج كافٍ للمستقبلات اللمسية للجلد - الضغط ، لبراعم التذوق في اللسان - مجموعة متنوعة من المواد الكيميائية ، لعضلات الهيكل العظمي - النبضات العصبية التي تتدفق إليها على طول الأعصاب الحركية. غير كافية تلك المحفزات لإدراك أن الخلية أو العضو المعين غير مكيَّف بشكل خاص. لذلك ، تنقبض العضلة عند تعرضها للحمض أو القلوي ، التيار الكهربائي ، التمدد المفاجئ ، الصدمة الميكانيكية ، الاحترار السريع ، إلخ. الخلايا أكثر حساسية لمحفزاتها الكافية من تلك غير الكافية. هذا تعبير عن تكيف وظيفي تم تطويره في عملية التطور.
الاهتياجيةيتم تكييف بعض الخلايا والأنسجة (العصبية والعضلية والغدية) بشكل خاص للقيام بردود فعل سريعة للتهيج. تسمى هذه الخلايا والأنسجة بالإثارة ، وتسمى قدرتها على الاستجابة للتهيج بالإثارة. مقياس الاستثارة هو الحد الأدنى من قوة الحافز الذي يسبب الإثارة. هذه
الحد الأدنى من قوة التهيج يسمى عتبة التهيج. كلما زاد الحد الأدنى من قوة التهيج المطلوبة لإحداث ردود فعل ، كلما زادت عتبة التهيج ، وانخفضت الاستثارة ، وعلى العكس ، كلما انخفضت عتبة التهيج ، زادت الاستثارة. فيما يتعلق بالمنبهات المختلفة ، يمكن أن تكون عتبة التهيج مختلفة. إن استثارة المستقبلات فيما يتعلق بالمنبهات الكافية عالية بشكل خاص ، على سبيل المثال ، لتهيج خلية شمية ، يكفي أن تعمل عليها جزيئات قليلة فقط من مادة معطرة.
الإثارةتتميز الخلايا المستثارة بشكل محدد من الاستجابة لعمل المنبهات: تحدث فيها عملية فسيولوجية تشبه الموجة - الإثارة هي تفاعل بيولوجي معقد يتجلى في مزيج من العمليات الفيزيائية والفيزيائية والكيميائية والتغيرات الوظيفية. علامة الإثارة الإلزامية هي حدوث تغيير في الحالة الكهربائية لغشاء الخلية السطحي. عند الإثارة ، تنتقل الخلايا من حالة الراحة الفسيولوجية إلى حالة النشاط الفسيولوجي المميزة لخلية معينة: تتقلص الألياف العضلية ، وتفرز الخلية الغدية سرًا .. في الخلية المستثارة ، يوجد فرق مستمر في الجهد الكهربائي بين السيتوبلازم والبيئة الخارجية ، أي على جانبي أغشية الخلايا السطحية. وبالتالي فإن الأخير مستقطب - سطحه الداخلي مشحون سلبًا فيما يتعلق بالسطح الخارجي. هذا الاختلاف في الجهد يسمى جهد الغشاء. سبب هذا الاختلاف في الجهد هو عدم المساواة في تركيز الأيونات داخل الخلية - في سيتوبلازمها وخارج الخلية - في سائل الأنسجة المحيطة: يحتوي السيتوبلازم على المزيد من أيونات البوتاسيوم وعدد أقل من أيونات الصوديوم مقارنة بسوائل الأنسجة. في حالة الراحة ، يكون غشاء الخلية منفذاً قليلاً لأيونات الصوديوم. عند الإثارة ، تزداد نفاذية الغشاء ، ويمرر أيونات الصوديوم موجبة الشحنة إلى الخلية ، مما يؤدي إلى انخفاض فرق جهد الغشاء (إزالة الاستقطاب من الغشاء) وحتى ظهور فرق محتمل للعلامة المعاكسة. يسمى التغيير في فرق الجهد الكهربائي أثناء الإثارة بجهد الفعل. التيار الكهربائي المتولد
عندما يتم توصيل نسيج متحمس بنسيج غير متحمس ، فإنه يسمى تيار العمل. الإثارة ، كما كانت ، عملية انفجارية ناتجة عن تغيير في نفاذية الغشاء تحت تأثير مادة مهيجة. هذا التغيير في البداية صغير نسبيًا ويصاحبه فقط إزالة استقطاب طفيفة ، وانخفاض طفيف في إمكانات الغشاء في الموقع حيث تم تطبيق التحفيز ، ولا ينتشر على طول النسيج المثير (وهذا ما يسمى بالإثارة المحلية). بعد الوصول إلى مستوى - عتبة - حرج ، فإن التغيير في فرق الجهد ينمو مثل الانهيار الجليدي وبسرعة - في العصب في بضعة أجزاء من عشرة آلاف من الثانية - يصل إلى الحد الأقصى. استعادة فرق الجهد الأولي - عودة استقطاب الغشاء - يحدث في البداية بسبب إطلاق أيونات البوتاسيوم من الخلية. بعد ذلك ، بفضل آلية فسيولوجية خاصة ، ما يسمى بمضخة الصوديوم والبوتاسيوم ، يتم استعادة عدم المساواة في تركيزات الأيونات بين السيتوبلازم والبيئة المحيطة بالخلية (تدخل أيونات البوتاسيوم إلى الخلية مرة أخرى ، وتتركها أيونات الصوديوم). تتطلب عملية الاسترداد هذه بعض إنفاق الطاقة ، والتي يتم توفيرها من خلال عمليات التمثيل الغذائي. السمة المميزة للخلية في لحظة الإثارة - خلال فترة إزالة الاستقطاب القصوى للغشاء - هي عدم قدرتها على الاستجابة لتهيج جديد. تسمى حالة عدم استثارة الخلية أثناء إثارة الانكسار. الإثارة عملية متموجة. بعد أن نشأ في خلية واحدة أو في قسم واحد منها ، على سبيل المثال ، في قسم واحد من الألياف العصبية ، ينتشر الإثارة ، ويمر إلى خلايا أخرى أو إلى أقسام أخرى من نفس الخلية. يرجع إجراء الإثارة إلى حقيقة أن جهد الفعل الذي نشأ في خلية واحدة أو في إحدى مناطقها يصبح مهيجًا يسبب إثارة المناطق المرغوبة. ينتقل الإثارة من خلية عصبية إلى أخرى أو من ألياف عصبية إلى عضلة أو خلية غدية كيميائيًا. في نهاية العصب ، يتم تكوين مركبات كيميائية - أجهزة إرسال النبضات العصبية (أستيل كولين ، نورإبينفرين ، إلخ) ، مما يتسبب في إثارة في الخلية المثيرة التي تقع عليها نهاية العصب. تسمى المرسلات الكيميائية للنبضات العصبية بالناقلات العصبية.
يرتبط علم وظائف الأعضاء ارتباطًا وثيقًا بالعديد من العلوم الأخرى ، حيث يعتمد في بحثه على بيانات بعض العلوم والوجود. بدوره ، أساس تنمية الآخرين. الفيزياء والكيمياء والفيزياء الحيوية والكيمياء الحيوية وعلم التشريح وعلم الأنسجة وعلم الأجنة وعلم الأحياء العام وعلم الوراثة وعلم التحكم الآلي - هذه ليست قائمة كاملة من التخصصات ، التي تعتبر معرفتها ذات أهمية كبيرة لدراسة وفهم العمليات الفسيولوجية التي تحدث في الجسم.
ومع ذلك ، فأنت تستعد لتصبح أطباء ، وتستعد لعلاج المرضى. وعلم وظائف الأعضاء يدرس وظائف طبيعية. جسم سليم - لهذا يطلق عليه "علم وظائف الأعضاء الطبيعي". لماذا يحتاج الطبيب إلى معرفة وظائف الكائن الحي السليم ، إذا كان في عمله سيواجه المرضى فقط؟ ربما يكفي فقط "علم وظائف الأعضاء المرضي" الذي تمت دراسته في السنة الثالثة؟ يعتقد العديد من الطلاب ذلك ، وفي النهاية اتضح أنهم ليس لديهم معرفة قوية بعلم وظائف الأعضاء الطبيعي أو المرضي أو التخصصات السريرية.
أعتقد أن لديك فكرة جيدة عن سبب حاجة الطبيب إلى معرفة قوانين وآليات الشخص السليم. هل ستصلح جهاز تلفزيون إذا كنت لا تعرف مبادئ تشغيله ، إذا كنت لا تعرف كيف يجب أن يعمل جهاز قابل للصيانة؟ هل ستجمع السيارة إذا كنت لا تعرف قوانين التفاعل بجميع أجزائها وآلياتها؟ والإنسان آلة بيولوجية شديدة التعقيد ، وإصلاحها مهمة صعبة للغاية وحساسة وتتطلب مهارات عالية! افهم الاختلالات التي تحدث في الجسم أثناء امراض عديدة، لا يمكن تحديد الخطوط العريضة للطرق الصحيحة لعلاجهم إلا بمعرفة جيدة بالعمليات الفسيولوجية التي تحدث في الجسم الطبيعي السليم.
لا تقتصر أهمية علم وظائف الأعضاء على دراسة المعلمات الفسيولوجية الطبيعية للكائن الحي. هذا وحده من شأنه أن يساعد الطبيب فقط على تحديد ما إذا كان الشخص لديه انحرافات عن القاعدة. لكن علم وظائف الأعضاء يزود الطبيب بآليات لتصحيح الوظائف المضطربة ، مما يسمح بما يسمى بالعلاج "الممرض" ، والذي يقضي على سبب تطور المرض ، وليس أعراضه. لا يمكن للطبيب الحديث أن يصف العلاج على أساس مبدأ "من الرأس ، من المعدة ، من البرد". في عملية دراسة المريض على أساس معرفة علم وظائف الأعضاء ، يجب عليه العثور على سبب المرض ، والاعتماد مرة أخرى على علم وظائف الأعضاء ، وإجراء مثل هذا التدخل ، والذي يؤدي فقط إلى الشفاء الإجباري.
تسمح معرفة الأساليب الفسيولوجية لدراسة الوظائف للطبيب باختيار ما هو مطلوب حاليًا ، ويساعد على تقييم نتائج هذه الدراسات بشكل صحيح ، ومن ترسانة كبيرة أدويةوالطرق الطبية لاختيار الأكثر فعالية في هذه الحالة.
تساهم دراسة المرض الذي يصيب الإنسان في فهم العديد من العمليات الفسيولوجية الطبيعية. ا. كتب بافلوف عن هذا: "إن عالم الظواهر المرضية هو سلسلة لا نهاية لها من كل التوليفات الممكنة ، الخاصة ، ... للظواهر الفسيولوجية التي لا تحدث عادة. إنها مثل سلسلة من التجارب الفسيولوجية التي أجرتها الطبيعة والحياة ، وهذا غالبًا ما يكون مزيجًا من الظواهر التي لم تكن لتحدث لعلماء وظائف الأعضاء المعاصرين ، والتي في بعض الأحيان لا يمكن حتى إعادة إنتاجها عن قصد الوسائل التقنيةعلم وظائف الأعضاء الحديث. تفرض العيادة مهامًا جديدة لعلم وظائف الأعضاء وفي نفس الوقت تعد مصدرًا غنيًا للأعمال الفسيولوجية الجديدة. إن أهمية علم وظائف الأعضاء للطب والطب لعلم وظائف الأعضاء كبيرة جدًا لدرجة أن اتحادهم الوثيق هو الوحيد الذي يمكنه إقامة صرح علمي معقد حول جسم الانسان.
علم وظائف الأعضاء الحديث هو علم تجريبي: يعتمد البحث الفسيولوجي على التجربة والتجربة. في التجربة ، يتم إجراء تداخل اصطناعي في عمليات معينة في الجسم من أجل توضيح دور وأهمية وترابط الأجهزة والأنظمة الفردية. لذلك ، على سبيل المثال ، عندما يتم تهيج العصب السمبثاوي العنقي بواسطة تيار كهربائي ، يتم تضييق أوعية الأذن على الجانب الذي يتم تشغيله ، ويؤدي قطع هذا العصب إلى توسع حاد في هذه الأوعية. أتاحت هذه التجارب لعالم وظائف الأعضاء كلود برنارد أن ينشئ لأول مرة التنظيم العصبي لتجويف الأوعية الدموية.
في ترسانة علم وظائف الأعضاء ، هناك طرق مثل الاستئصال ، والزرع ، وإزالة التعصيب ، وطريقة تطبيق الأربطة والمفاغرة ، وتقنيات الناسور ، وطرق التهيج والقطع ، والتروية وتسجيل الوظائف ، إلخ. تتطلب معظم هذه التقنيات تشريح جثة الحيوان أو الجراحة. يتم استخدامها في التجارب الحادة أو المزمنة.
لا يمكن استخدام ترسانة طرق البحث المستخدمة في التجارب على الحيوانات في الدراسة جسم الانسان. لذلك ، كانت المعلومات حول وظائف العديد من الأعضاء لدى البشر ، حتى وقت قريب ، تقتصر إلى حد كبير على البيانات التي تم الحصول عليها في التجارب على الحيوانات. في الوقت الحاضر ، تغير الوضع من نواح كثيرة. وضعت إنجازات الفيزياء الحديثة وهندسة الراديو والإلكترونيات وعلم التحكم الآلي في أيدي علماء وظائف الأعضاء مثل هذه الأجهزة التي تسمح ، دون التدخل في جسم الإنسان ، بتسجيل عمل أعضائه. سترى العديد من هذه الأجهزة أثناء دراستك في قسم علم وظائف الأعضاء الطبيعي.
الأنماط العامة لتنظيم الوظائف.
موضوع علم وظائف الأعضاء هو في المقام الأول وظائف الجسم وأجزائه. لذلك ، بالانتقال إلى عرض علم وظائف الأعضاء ، يجب علينا أولاً وقبل كل شيء أن نتناول مفاهيم التنظيم والوظيفة.
إن ORGANISM هي وحدة موجودة بشكل مستقل في العالم العضوي ، وهي عبارة عن نظام ذاتي التنظيم يتفاعل مع التغيرات المختلفة في البيئة الخارجية ككل ، وقادر على الوجود فقط من خلال التفاعل المستمر مع البيئة الخارجية والتجديد الذاتي كنتيجة لذلك. من هذا التفاعل.
الوظائف الفسيولوجية هي مظاهر النشاط الحيوي التي لها قيمة تكيفية وتهدف إلى تحقيق نتيجة معينة مفيدة للجسم. عند القيام بوظائف مختلفة ، يتكيف الجسم مع البيئة الخارجية أو يكيّف البيئة مع احتياجاته.
الوظيفة الرئيسية للكائن الحي هي التمثيل الغذائي والطاقة (التمثيل الغذائي). تتكون هذه العملية من مزيج من المواد الكيميائية و تغيرات فيزيائية، في تحول المواد والطاقة ، بشكل مستمر ومستمر في الجسم وفي جميع هياكله. التمثيل الغذائي هو شرط ضروري للحياة. هو الذي يميز الحي عن غير الحي. الحياة ممكنة فقط طالما يوجد التمثيل الغذائي الذي يحافظ على وجود البروتوبلازم الحي وتجديده الذاتي. يؤدي توقف التمثيل الغذائي إلى تدمير البروتوبلازم وموت الكائن الحي.
ترتبط جميع الوظائف الفسيولوجية الأخرى بعملية التمثيل الغذائي ، سواء كانت النمو ، أو التطور ، أو التكاثر ، أو التغذية ، أو الهضم ، أو التنفس ، أو الإخراج ، أو الحركة ، أو ردود الفعل على التغيرات في البيئة الخارجية. يهدف نشاطهم في المقام الأول إلى الحفاظ على ظروف التمثيل الغذائي المثلى. (ضمان التشغيل الطبيعي لما يسمى بغلاية التمثيل الغذائي). في نفس الوقت ، أساس أي وظيفة هو مجموعة معينة من تحولات المادة والطاقة. وهذا ينطبق بالتساوي على وظائف خلية واحدة ، أو نسيج ، أو عضو ، أو كائن حي ككل.
يحتاج كل كائن حي ، أحادي الخلية أو متعدد الخلايا ، إلى ظروف وجود معينة يوفرها له ذلك الموطن (ما يسمى بالبيئة الخارجية) التي تكيف معها هذا النوع من الكائنات الحية طوال المسار الكامل لتطوره التطوري. عادة ما يتم توفير وظائف الجسم فقط إذا أعطته البيئة الخارجية الفرصة لتلقي الطعام ، عند درجة حرارة معينة ، وضغط بارومتري ، وشدة وطيف من الضوء ، وما إلى ذلك.
في الوقت نفسه ، يجب ألا يغيب عن البال أن حدود التقلبات البيئية التي يتحملها الكائن الحي للحيوانات الأعلى أوسع بكثير من تلك اللازمة للتشغيل الطبيعي لمعظم خلاياه. والسبب في ذلك أن موطن خلايا الجسم هو بيئته الداخلية ، والتي تتغير بدرجة أقل بكثير من البيئة الخارجية. البيئة الداخلية للجسم هي الدم والليمفاوية وسوائل الأنسجة التي تعيش فيها الخلايا.
وظائف خلايا الجسم طبيعية فقط بثبات نسبي
الضغط الاسموزي ، وتركيب المنحل بالكهرباء ، وتركيز معين من أيونات الهيدروجين ، والموارد الغذائية والطاقة. ثبات التركيب الكيميائي والخصائص الفيزيائية والكيميائية للبيئة الداخلية ميزة مهمةكائنات حيوانية أعلى. للدلالة على هذا الثبات ، اقترح كانون المصطلح التوازن. التعبير عن التوازن هو وجود عدد من العوامل البيولوجية الثوابت، بمعنى آخر. مؤشرات كمية مستقرة تميز الحالة الطبيعية للكائن الحي. اعتمادًا على قيمتها ، هناك ثوابت صلبة ولينة (بلاستيكية). مع ملاحظة ثبات التركيب والخصائص الفيزيائية والكيميائية والبيولوجية للبيئة الداخلية ، يجب التأكيد على أنها ليست مطلقة ، ولكنها نسبية وديناميكية. يتحقق هذا الثبات من خلال العمل المستمر لعدد من الأجهزة والأنظمة ، ونتيجة لذلك فإن التحولات في التركيب والخصائص الفيزيائية والكيميائية للبيئة الداخلية التي تحدث تحت تأثير التغيرات في البيئة الخارجية والنشاط الحيوي لل يتم محاذاة الكائن الحي.
التوازن له حدود معينة. أثناء الإقامة الطويلة بشكل خاص ، في ظل ظروف تختلف اختلافًا كبيرًا عن تلك التي يتكيف معها الجسم ، يحدث اضطراب الاستتباب ويمكن أن تحدث تحولات غير متوافقة مع الحياة. حتى الانتهاكات الصغيرة للتوازن تؤدي إلى علم الأمراض ، وبالتالي ، فإن تحديد الثوابت الفسيولوجية الثابتة نسبيًا (الرقم الهيدروجيني ، وضغط الدم ، ومعدل التنفس ، ومعدل ضربات القلب ، و MOD ، وما إلى ذلك) له قيمة تشخيصية كبيرة. لذلك ، يتم تنفيذ عمل الحفاظ على التوازن باستمرار وبشكل مستمر من خلال الأنظمة التنظيمية ذات الصلة ، والتي سنتحدث عنها لاحقًا.
يختلف دور الأجهزة المختلفة وأنظمتها في الحفاظ على التوازن. سنتحدث عنها أيضًا في الأقسام ذات الصلة من الدورة.
كما ذكرنا سابقًا ، فإن السمة المميزة لأي كائن حي هي أنه نظام ذاتي التنظيم يستجيب للتأثيرات المختلفة ككل. مبدأ التنظيم الذاتي هو أن انحراف أي ثابت عن المستوى الطبيعي هو بحد ذاته إشارة لتصحيح هذه التحولات. يتحقق التنظيم الذاتي من خلال تفاعل جميع خلايا الجسم وأنسجته وأعضائه. يتم التعبير عن هذا التفاعل بين الأجهزة بشكل خاص في عمل ما يسمى. أنظمة وظيفية. يتكون هذا النظام من أجهزة ، يضمن نشاطها المشترك التكيف مع ظروف بيئية معينة ، مما يضمن تلبية أي حاجة داخلية.
تحت الاحتياجات الداخليةمن الآن فصاعدًا ، سوف نفهم أي انحراف طويل المدى أكثر أو أقل لواحد أو آخر من ثابت بيئته الداخلية عن المستوى الذي يضمن نشاط حياته الطبيعي. إن الاحتياجات البيولوجية هي الدافع الأول في سلسلة عمليات التنظيم الذاتي لوظائف الجسم المختلفة.
يمكن أن تكون احتياجات الكائن الحي لا حصر لها. ومع ذلك ، فإنهم جميعًا يتحدون في مجموعات كبيرة - بيولوجية ، واجتماعية ، وجنسية ، وغذائية ، ودفاعية ، وما إلى ذلك. إرضاء واحد أو آخر من الاحتياجات ويمثل لكل كائن حي شيئًا معينًا نتيجة مفيدةنشاطه التكيفي ، أي المهام.
التنظيم هو تغيير موجه في وظائف الأعضاء والأنسجة المختلفة. يتم تنظيم جميع وظائف الهيئة من خلال آليتين تنظيميتين رئيسيتين - عصبي وخلطي.
آلية خلطيةيعتمد على حقيقة أنه في الخلايا والأعضاء المختلفة ، أثناء عملية التمثيل الغذائي ، يتم تشكيل مواد كيميائية ذات طبيعة مختلفة وعمل فسيولوجي. يدخل سائل الأنسجة ، ثم إلى الدم ، وينتقل إلى جميع أنحاء الجسم ، ويمكن أن يؤثر على خلايا الأنسجة. حالة خاصة من التنظيم الخلطي هي الغدد الصماء ، التي تقوم بها الغدد الصماء.
الآلية العصبيةيكمن التنظيم في حقيقة أن أوامر البدء أو التعديل يتم إرسالها عبر الأعصاب إلى جميع الخلايا والأعضاء ، وتغيير نشاطها في الاتجاه اللازم للجسم. التغيرات في حالة بعض الخلايا والأعضاء من خلال الجهاز العصبي بطريقة انعكاسية تسبب تغيرات في وظائف الأعضاء الأخرى. تعد آلية التنظيم هذه أكثر كمالًا ، نظرًا لأن تفاعل الخلايا من خلال الجهاز العصبي يتم بشكل أسرع بكثير من التفاعل الكيميائي الخلطي ، وبالإضافة إلى ذلك ، فإن النبضات العصبية دائمًا ما يكون لها عنوان محدد.
وهكذا ، اثنان مبادئ عامةتنظيم جميع وظائف الجسم:
1. جميع وظائف الجسم ينظمها الجهاز العصبي و / أو الخلطي.
2. يتم تنظيم الوظائف وفقًا لمبدأ التنظيم الذاتي.
يتجلى كلا المبدأين بشكل واضح وكامل في أنشطة ما يسمى أنظمة وظيفية(FS) ، والتي تتشكل باستمرار عند ظهور حاجة في الجسم وتضمن إشباعها الأمثل.
النظام الوظيفي هو مجموعة من الأعضاء والأنسجة غير المتجانسة التي تتحد على أساس وظيفي وتوفر ، عند التفاعل ، وظائف وأشكال نشاط جديدة نوعياً ، مع نتيجة متأصلة في النظام ككل وليست متأصلة في أجزائه بشكل منفصل. FS هي منظمة ديناميكية ذاتية التنظيم ، أنشطة الجميع العناصر المكونةمما يساهم في الحصول على نتيجة تكيفية حيوية للجسم.
يمكن أن يشمل تكوين FS مجموعة متنوعة من الأعضاء والأنسجة ، والتي يمكن أن يؤدي نشاطها إلى استعادة التوازن المضطرب. لا يتم تنظيم الأنظمة الوظيفية وفقًا للصفات التشريحية ، ولكن وفقًا للخصائص الفسيولوجية. العامل الرئيسي لتشكيل النظام هو الهدف ، نتيجة الأنشطة المستقبلية للخدمة الثابتة. مع تقدمنا في دورة علم وظائف الأعضاء ، سنقوم بتمييز العديد من FS (الحفاظ على الرقم الهيدروجيني ، والضغط الاسموزي ، وتركيز المغذيات في ضغط الدم ، وما إلى ذلك) الآن يجب علينا النظر في المخطط العام لهيكل أي FS. هذا المخطط العام ، التنظيمي المحدد والآليات التنفيذية ونتائج أنشطتها ، لبناء مخطط أي FS.
عامل تشكيل النظام المركزي لكل PS هو نتيجة لعمله ، والذي يحدد الظروف الطبيعية لمسار عمليات التمثيل الغذائي في الجسم ككل. ويترتب على ذلك أن النتيجة هي نوع من "بطاقة الاتصال" لكل FS.
في الكائن الحي ، يمكن تمييز 3 مجموعات من النتائج التكيفية المفيدة.
تتكون المجموعة الأولى من الثوابت الداخلية للجسم ، والمؤشرات الاستتبابية التي تحدد نشاطه الحيوي الطبيعي: درجة الحموضة ، وتركيز الأملاح ، والمغذيات ، والغازات ، إلخ.
تتكون المجموعة الثانية من نتائج النشاط التكيفي للكائن الحي في البيئة ، والذي يهدف إلى تلبية احتياجاته البيولوجية الداخلية ، والحفاظ على النوع والجنس (السلوك الهادف الذي يؤدي إلى إخماد العطش والجوع ، وما إلى ذلك).
المجموعة الثالثة هي نتائج النشاط الاجتماعي للشخص الذي يهدف إلى تلبية احتياجاته الاجتماعية.
اعتمادًا على خصائص النتيجة ، ستكون FS أبسط أو أكثر تعقيدًا ، لكن مخططها العام يظل كما هو. هذا الأخير ، من خلال الأجهزة التنفيذية ، يعيد مرة أخرى النتيجة التكيفية المفيدة إلى المستوى المطلوب. تتواصل كل هذه العمليات باستمرار مع إعلام المركز باستمرار بنجاح تحقيق نتيجة تكيفية مفيدة. يمكن تعبئة نفس المشغلات والأجهزة الطرفية لأداء وظائف الجسم المختلفة وتكون جزءًا من FS مختلفة.
الكائن الحي هو أكثر الآليات تعقيدًا ، ويتألف من العديد من FSs التي تتلاءم بشكل وثيق في عملها ولديها النقاط المشتركةاتصال وتسلسل هرمي معين. ومع ذلك ، في كل حالة محددة ، في كل لحظة زمنية محددة ، هناك دائمًا FS مهيمن ، والذي يحدد نشاط الكائن الحي في لحظة معينة ويخضع نشاط الأنظمة الوظيفية الأخرى.
يتحد عمل العديد من FS في الجسم بالدم ، كنقطة تركيز للنتائج النهائية للعديد من FS. بما أن الدم هو البيئة الداخلية للجسم ، وبما أن ثوابته متوازنة وصلبة إلى حد كبير ، فإننا نبدأ دراسة موضوع علم وظائف الأعضاء في قسم "فسيولوجيا نظام الدم".
محاضراتفي الفسيولوجيا البشرية
علم وظائف الأعضاء كعلم. الموضوع والمهام والأساليب وتاريخ علم وظائف الأعضاء
علم وظائف الأعضاء (physis - الطبيعة) هو علم عمليات الحياة الطبيعية للجسم وأنظمته الفسيولوجية المكونة له والأعضاء الفردية والأنسجة والخلايا والهياكل دون الخلوية وآليات تنظيم هذه العمليات وتأثير العوامل البيئية الطبيعية على الجسم. المهام.
بناءً على هذا ، بشكل عام ، فإن موضوع علم وظائف الأعضاء هو كائن حي صحي. يتم تضمين مهام علم وظائف الأعضاء في تعريفه. الطريقة الرئيسية لعلم وظائف الأعضاء هي التجربة على الحيوانات. هناك نوعان رئيسيان من التجارب أو التجارب:
1. تجربة حادة أو تشريح (قطع حي). في أثناء ذلك ، يتم إجراء تدخل جراحي ، ويتم فحص وظائف العضو المفتوح أو المعزول. بعد ذلك ، لا يسعون إلى بقاء الحيوان. تتراوح مدة التجربة الحادة من عدة عشرات من الدقائق إلى عدة ساعات (مثال).
2. التجربة المزمنة. في سياق التجارب المزمنة ، يتم إجراء التدخل الجراحي للوصول إلى العضو. ثم يحققون التئام الجروح الجراحية وبعد ذلك فقط يبدؤون البحث. يمكن أن تكون مدة التجارب المزمنة عدة سنوات (مثال).
في بعض الأحيان يتم تمييز تجربة تحت الحاد (مثال).
في الوقت نفسه ، يتطلب الطب معلومات حول آليات عمل جسم الإنسان. لذلك ، فإن I.P. كتب بافلوف: "لا يمكن تطبيق البيانات التجريبية على الشخص إلا بحذر ، والتحقق باستمرار من واقعية أوجه التشابه مع نشاط هذه الأعضاء في الإنسان والحيوان." وبالتالي ، بدون ملاحظات وتجارب خاصة على الشخص ، فإن دراسة له علم وظائف الأعضاء لا معنى له ، لذلك يتميز علم فسيولوجي خاص - فسيولوجيا الإنسان ، وعلم وظائف الأعضاء البشري له موضوع ومهام وطرق وتاريخ ، وموضوع فسيولوجيا الإنسان هو جسم الإنسان السليم.
مهامها:
1. دراسة آليات عمل الخلايا والأنسجة والأعضاء وأنظمة جسم الإنسان ككل
2. دراسة آليات تنظيم وظائف أجهزة وأنظمة الجسم.
3. الكشف عن تفاعلات جسم الإنسان وأنظمته مع التغيرات في البيئة الخارجية والداخلية.
نظرًا لأن علم وظائف الأعضاء ككل هو علم تجريبي ، فإن الطريقة الرئيسية لفيزيولوجيا الإنسان هي أيضًا التجربة. ومع ذلك ، فإن التجارب على البشر تختلف اختلافًا جوهريًا عن التجارب على الحيوانات. أولاً ، يتم إجراء الغالبية العظمى من الأبحاث البشرية باستخدام طرق غير جراحية ، أي بدون تدخل في الأعضاء والأنسجة (مثال على تخطيط القلب ، تخطيط كهربية القلب ، تخطيط كهربية القلب ، اختبارات الدم ، إلخ). ثانياً ، لا يتم إجراء التجارب على البشر إلا عندما لا تضر بالصحة وبموافقة الشخص المعني. في بعض الأحيان يتم إجراء تجارب حادة على شخص في عيادة عندما تتطلب مهام التشخيص ذلك (مثال). ومع ذلك ، تجدر الإشارة إلى أنه بدون بيانات علم وظائف الأعضاء الكلاسيكي ، كان ظهور وتطور علم وظائف الأعضاء البشري مستحيلًا (آثار لضفدع وكلب). المزيد كتب بافلوف ، في تقييمه لدور علم وظائف الأعضاء في الطب ، ما يلي: "يُفهم بالمعنى التقريبي للكلمة ، علم وظائف الأعضاء والطب لا ينفصلان ، ومعرفة علم وظائف الأعضاء ضرورية لأي طبيب في أي تخصص." وأيضًا أن "الطب ، الذي يتم إثراؤه باستمرار يومًا بعد يوم ، بحقائق فسيولوجية جديدة ، سيصبح أخيرًا ما ينبغي أن يكون عليه بشكل مثالي ، أي القدرة على إصلاح آلية بشرية مدللة وتطبيق المعرفة في علم وظائف الأعضاء" (أمثلة من العيادة) . عالم فسيولوجي روسي مشهور آخر البروفيسور. في يا. وأشار Danilevsky: "كلما تم تحديد العلامات المعيارية للحياة الجسدية والعقلية للشخص بشكل أكثر دقة وكاملة ، كلما كان تشخيص الطبيب أكثر صحة لتشوهاته المرضية."
علم وظائف الأعضاء ، لكونه علمًا بيولوجيًا أساسيًا ، يرتبط ارتباطًا وثيقًا بالأساسيات الأخرى و العلوم البيولوجية. على وجه الخصوص ، بدون معرفة قوانين الفيزياء ، من المستحيل شرح الظواهر الكهربية الحيوية ، آليات إدراك الضوء والصوت. بدون استخدام بيانات الكيمياء ، من المستحيل وصف عمليات التمثيل الغذائي ، والهضم ، والتنفس ، وما إلى ذلك. لذلك ، على حدود هذه العلوم مع علم وظائف الأعضاء ، تبرز العلوم البنت للفيزياء الحيوية والكيمياء الحيوية.
نظرًا لأن الهيكل والوظيفة لا ينفصلان ، وهي الوظيفة التي تحدد تكوين الهيكل ، فإن علم وظائف الأعضاء وثيق الصلة بالعلوم المورفولوجية: علم الخلايا ، وعلم الأنسجة ، وعلم التشريح.
نتيجة لدراسة تأثير المواد الكيميائية المختلفة على الجسم ، ظهر علم العقاقير والسموم من علم وظائف الأعضاء إلى العلوم المستقلة. كان تراكم البيانات حول انتهاكات آليات عمل الجسم في الأمراض المختلفة بمثابة أساس لظهور علم وظائف الأعضاء المرضي.
يميز بين علم وظائف الأعضاء العام والخاص. يدرس علم وظائف الأعضاء العام الأنماط الأساسية لحياة الجسم ، وآليات العمليات الأساسية مثل التمثيل الغذائي والطاقة ، والتكاثر ، وعمليات الإثارة ، إلخ. يدرس علم وظائف الأعضاء الخاص وظائف خلايا وأنسجة وأعضاء وأنظمة فسيولوجية معينة. لذلك ، فإنه يسلط الضوء على أقسام مثل فسيولوجيا الأنسجة العضلية ، والقلب ، والكلى ، والهضم ، والتنفس ، وما إلى ذلك. بالإضافة إلى ذلك ، يوجد في علم وظائف الأعضاء أقسام لها موضوع معين للدراسة أو مناهج خاصة لدراسة الوظائف. وتشمل هذه علم وظائف الأعضاء التطوري (التفسير) ، وعلم وظائف الأعضاء المقارن ، وعلم وظائف الأعضاء العمر.
هناك عدد من الفروع التطبيقية في علم وظائف الأعضاء. هذا ، على سبيل المثال ، فسيولوجيا حيوانات المزرعة. في علم وظائف الأعضاء البشرية ، تتميز الأقسام التطبيقية التالية:
1. علم وظائف الأعضاء العمر. يدرس السمات المرتبطة بالعمر لوظائف الجسم.
2. فسيولوجيا العمل.
3. علم وظائف الأعضاء السريرية. هذا علم يستخدم الأساليب والنهج الفسيولوجية لتشخيص وتحليل التشوهات المرضية.
4. علم وظائف الطيران والفضاء.
5. فسيولوجيا الرياضة.
يرتبط علم وظائف الأعضاء البشري ارتباطًا وثيقًا بالتخصصات السريرية مثل العلاج والجراحة والتوليد وعلم الغدد الصماء والطب النفسي وطب العيون وما إلى ذلك. على سبيل المثال ، تستخدم هذه العلوم طرقًا عديدة طورها علماء وظائف الأعضاء للتشخيص. أساس الكشف عن علم الأمراض هو الانحرافات في المعايير الطبيعية للجسم.
بعض أقسام علم وظائف الأعضاء البشرية هي أساس علم النفس. هذا هو فسيولوجيا الجهاز العصبي المركزي ، والنشاط العصبي العالي ، أنظمة حسية، علم النفس الفسيولوجي.
تم وصف تاريخ علم وظائف الأعضاء بالتفصيل في الكتاب المدرسي ، محرر. تكاتشينكو
^ آليات تنظيم وظائف الجسم
التنظيم الخلطي والعصبي. لا ارادي. القوس الانعكاسي. المبادئ الأساسية لنظرية الانعكاس
يتم تنظيم جميع وظائف الجسم من خلال نظامين من التنظيم: خلطي وعصبي. التنظيم الخلطي الأقدم نسبيًا هو التنظيم عن طريق المواد الفعالة فيزيولوجيًا (PAS) المنتشرة في سوائل الجسم - الدم ، اللمف ، السائل بين الخلايا. عوامل التنظيم الخلطي هي:
1. المستقلبات والأيونات غير العضوية. على سبيل المثال ، كاتيونات الكالسيوم والهيدروجين وثاني أكسيد الكربون.
2. هرمونات الغدد الصماء. تنتجها غدد صماء متخصصة. هذه هي الأنسولين ، هرمون الغدة الدرقية ، إلخ.
3-الهرمونات المحلية أو الأنسجة. يتم إنتاج هذه الهرمونات بواسطة خلايا خاصة تسمى paracrine ، ويتم نقلها بواسطة سائل الأنسجة وتعمل فقط على مسافة قصيرة من الخلايا المفرزة. وتشمل هذه المواد مثل الهيستامين والسيروتونين والهرمونات المعدية المعوية وغيرها.
4. المواد الفعالة بيولوجيا التي توفر روابط إبداعية بين خلايا الأنسجة. هذه هي جزيئات البروتين التي تفرزها. إنها تنظم التمايز والنمو والتطور لجميع الخلايا التي تتكون منها الأنسجة وتوفر ارتباطًا وظيفيًا للخلايا في الأنسجة. مثل هذه البروتينات هي ، على سبيل المثال ، keyons ، التي تمنع تخليق الحمض النووي وانقسام الخلايا.
السمات الرئيسية للتنظيم الخلطي:
1- انخفاض سرعة الإجراء التنظيمي المرتبط بالسرعة المنخفضة لتيارات سوائل الجسم المقابلة.
2. زيادة بطيئة في قوة الإشارة الخلطية وانخفاض بطيء. ويرجع ذلك إلى الزيادة التدريجية في تركيز PAS وتدميرها التدريجي.
3. عدم وجود نسيج معين أو عضو مستهدف لعمل العوامل الخلطية. تعمل على جميع الأنسجة والأعضاء على طول تدفق السوائل ، في الخلايا التي توجد بها مستقبلات مقابلة.
التنظيم العصبي هو تنظيم وظائف الجسم من خلال ردود الفعل التي يقوم بها الجهاز العصبي.
تم تطوير مفهوم مبدأ رد الفعل لنشاط الجهاز العصبي لأول مرة في القرن السابع عشر من قبل عالم الطبيعة الفرنسي رينيه ديكارت. اقترح مخططًا افتراضيًا لتشكيل الحركة اللاإرادية (التمثيل الآلي). تم إدخال مصطلح "منعكس" (عمل انعكاسي) في علم وظائف الأعضاء في عام 1771 بواسطة Unzer. طور J. Prohaska في عام 1800 مخططًا لأبسط قوس انعكاسي. هم. قام Sechenov بتوسيع مفهوم "المنعكس" ليشمل أي نشاط عصبي أعلى (HNA). في الوقت نفسه ، انطلق من فقرتين: 1. يتم اختزال أي نشاط للكائن الحي في النهاية إلى حركة. 2. جميع الحركات هي ردود فعل في الأصل. ا. أثبت بافلوف تجريبياً وجهة نظر المنعكس كالفعل الرئيسي لأي نشاط عصبي. كما قام أيضًا بتقسيم جميع ردود الفعل ، وفقًا لآلية التكوين ، إلى غير مشروطة وشرطية. الملامح الرئيسية لنظرية رد الفعل لـ I.P. صاغ بافلوف في عمله "إجابة عالم وظائف الأعضاء لعلماء النفس". وهو يتضمن ثلاثة مبادئ أساسية:
1. مبدأ الحتمية. تقول "لا عمل بدون سبب". أولئك. كل فعل منعكس هو نتيجة لعمل التحفيز على الكائن الحي.
2. مبدأ التحليل والتركيب. التحليل يحدث باستمرار في الدماغ. تمييز الإشارة ، وكذلك التوليف ، أي تفاعلهم وتصورهم الشامل.
3. مبدأ الهيكل. في الجهاز العصبيلا توجد عمليات ليس لها توطين هيكلي معين.
الأساس الصرفيأي منعكس هو قوس منعكس أو مسار انعكاسي. القوس الانعكاسي (RD) هو مسار التفاعل الانعكاسي ، أي إشارات عصبية. يتكون القوس الانعكاسي للانعكاس الجسدي (المحرك) من الروابط الرئيسية التالية:
1. مستقبلات تدرك التهيج
2. ألياف عصبية واردة أو صاعدة أو حسية
3. مركز العصب في C.N.S.
4. ألياف عصبية حركية فعالة أو تنازلية
5- الهيئة التنفيذية "الجهة المنفّذة"
يوجد في عدد من أقواس المنعكسات عصبون تغذية مرتدة (6) ، أو عصبون توكيد عكسي ، يتفاعل مع الاستجابة الانعكاسية ويتحكم فيها.
في القوس الانعكاسي الجسدي ، يمكن تمييز الخلايا العصبية التي تؤدي وظائف معينة. على وجه الخصوص ، في أبسط انعكاس أحادي المشبك لا يوجد سوى 2 من الخلايا العصبية: حساسة والمحرك. في أبسط قوس منعكس متعدد المشابك الذي نفكر فيه ، هناك: أ) خلية عصبية حساسة ، ب) خلية عصبية بينية ، ج) خلية عصبية تنفيذية. أرز. هناك المئات والآلاف من الخلايا العصبية في أقواس منعكسة معقدة متعددة المشابك.
في قوس الانعكاس اللاإرادي ، توجد الروابط التالية:
1. مستقبلات
2. وارد الألياف العصبية.
3. مركز عصبي (على سبيل المثال) لردود الفعل الودية في القرون الجانبية الحبل الشوكي)
4. الألياف العصبية Preganglionic
5. العقدة اللاإرادية
6- الألياف العصبية ما بعد العقدة
7. الهيئة التنفيذية. أرز.
النظم البيولوجية والوظيفية
في الخمسينيات والستينيات من القرن الماضي ، طور عالم الأحياء الكندي لودفيج برتالانفي ، باستخدام المناهج الرياضية والسيبرانية ، المبادئ الأساسية لتشغيل النظم البيولوجية. وتشمل هذه:
1. النزاهة ، أي عدم قابلية اختزال خصائص النظام إلى مجموع بسيط لخصائص أجزائه. أولئك. من المستحيل وصف خصائص النظام البيولوجي من خلال وظائف عناصره الفردية (مثال).
2. الهيكلية. إمكانية شرح وظائف النظام من خلال هيكله (مثال).
3. التسلسل الهرمي ، وإخضاع عناصر النظام لبعضها البعض من الأعلى إلى الأسفل. أولئك. المكونات العلوية للنظام تدير المكونات الأساسية (مثال).
4. العلاقة بين النظام والبيئة (مثال).
ومع ذلك ، لم يكشف برتالانفي عن العامل الأكثر أهمية - عامل تشكيل النظام. لذلك ، فإن الدور الرئيسي في تحديد الأنماط الجهازية المتأصلة في الكائنات الحية يعود إلى الأكاديمي P.K. أنوخين. في علم وظائف الأعضاء ، كان مفهوم النظم الفسيولوجية موجودًا منذ فترة طويلة. هذا عبارة عن مجموعة من الأعضاء الموحدة شكليًا ووظيفيًا والتي لها آليات تنظيمية مشتركة وتؤدي وظائف متجانسة (مثال). ومع ذلك ، P.K. أثبت أنوخين أن هناك أجهزة أخرى في الجسم ، على سبيل المثال ، توفر الدعم الحيوي. معلمات الجسم الهامة. أطلق عليها أنظمة وظيفية (FUS). وفقًا لـ P.K. Anokhin FUS عبارة عن مجموعة من الأعضاء والأنسجة التي تضمن تحقيق هدف في نوع معين من نشاط الحياة. يسمى هذا الهدف نتيجة تكيفية مفيدة (PPR). يمكن أن تكون أي معلمة للبيئة الداخلية ، على سبيل المثال ، درجة حرارة الجسم ، ومحتوى الأكسجين الطبيعي في الدم ، وما إلى ذلك ، نتيجة السلوك الذي يرضي بيولوجيًا ، على سبيل المثال ، الحاجة الغذائية ، نتيجة النشاط الاجتماعي للشخص. من المهم للطبيب أن يفهم FUS التي توفر التوازن.
إن طاعون المجترات الصغيرة هو العامل الذي يوحد مختلف أجهزة وأنظمة الجسم في كل واحد - FUS. لا يحدث ارتباط الأعضاء في FUS وفقًا للمورفولوجية ، ولكن وفقًا للخاصية الوظيفية. لذلك ، يمكن أن تشمل FUS أعضاء وأنسجة من مجموعة متنوعة من الأنظمة الفسيولوجية. علاوة على ذلك ، يمكن تضمين نفس الأعضاء في عدة فتحات في آن واحد. بالإضافة إلى ذلك ، على عكس الأنظمة الفسيولوجية ، يمكن توريث FUS وتشكيله في نفس الوقت الحياة الفردية. يتضمن مخطط FUS العام للحفاظ على معلمات التوازن العناصر التالية:
2. مستقبلات طاعون المجترات الصغيرة
3. وارد المسار
4. مركز العصب
5. التنظيم الخضري
6. التنظيم الخلطي
7. تنظيم السلوك
8. التمثيل الغذائي (الشكل)
إذا تجاوز طاعون المجترات الصغيرة ، تحت تأثير أي أسباب ، حدود القاعدة الفسيولوجية ، تكون مستقبلات طاعون المجترات الصغيرة متحمسة. تأتي النبضات العصبية منها إلى مركز العصب الذي ينظم هذه الوظيفة. من ذلك يذهبون إلى الأجهزة التنفيذية التي تضمن الحفاظ على معامل التوازن المناسب. في الوقت نفسه ، يتم إطلاق الآليات الأخلاقية للتنظيم. عندما لا يصل طاعون المجترات الصغيرة ، على الرغم من ذلك ، إلى المستوى الأولي ، تبدأ النبضات العصبية من مركز العصب بالتدفق إلى القشرة الدماغية. نتيجة لإثارة الخلايا العصبية ، يتم تشغيل الرابط الخارجي للتنظيم الذاتي للجسم ، أي تنظيم السلوك. هذا تغيير هادف في سلوك كائن حي. نتيجة لهذه الإجراءات التنظيمية ، يأتي تقرير أداء البرنامج إلى المستوى الأولي ، أي القاعدة الفسيولوجية. يتأثر طاعون المجترات الصغيرة بشكل مباشر بعملية التمثيل الغذائي. من ناحية أخرى ، فإن PPR نفسها لها تأثير مباشر على التمثيل الغذائي العمليات. أمثلة على عمل فتحات تسليك مختلفة.
^ مبادئ التنظيم الذاتي للجسم. مفهوم الاستتباب
والحركة المنزلية
القدرة على التنظيم الذاتي هي الخاصية الأساسية للأنظمة الحية ، ومن الضروري تهيئة الظروف المثلى لتفاعل جميع العناصر التي يتكون منها الجسم ، لضمان سلامته. هناك أربعة مبادئ رئيسية للتنظيم الذاتي:
1. مبدأ عدم التوازن أو التدرج. يكمن الجوهر البيولوجي للحياة في قدرة الكائنات الحية على الحفاظ على حالة ديناميكية غير توازن ، بالنسبة للبيئة. على سبيل المثال ، درجة حرارة جسم الحيوانات ذوات الدم الحار أعلى أو أقل من درجة حرارة البيئة. يوجد المزيد من كاتيونات البوتاسيوم في الخلية ، والصوديوم خارجه ، وما إلى ذلك. يتم ضمان الحفاظ على المستوى الضروري من عدم التناسق فيما يتعلق بالبيئة من خلال عمليات التنظيم.
2. مبدأ حلقة التحكم المغلقة. لا يستجيب كل نظام حي لمحفز فحسب ، بل يقيم أيضًا استجابة الاستجابة للحافز الحالي. أولئك. كلما كان التهيج أقوى ، زادت الاستجابة والعكس صحيح. يتم تنفيذ هذا التنظيم الذاتي بسبب العكس الإيجابي والسلبي استجابةفي أنظمة التنظيم العصبية والخلطية. أولئك. دائرة التنظيم مغلقة في حلقة. مثال على هذا الاتصال هو خلية عصبية واردة في أقواس الانعكاس الحركي.
3. مبدأ التنبؤ. الأنظمة البيولوجية قادرة على توقع نتائج الاستجابات بناءً على الخبرة السابقة. مثال على ذلك هو تجنب المنبهات المؤلمة بعد المنبهات السابقة.
4. مبدأ النزاهة. من أجل الأداء الطبيعي للنظام الحي ، يلزم سلامته الهيكلية.
تم تطوير عقيدة التوازن من قبل C. Bernard. في عام 1878 ، صاغ فرضية حول الثبات النسبي للبيئة الداخلية للكائنات الحية. في عام 1929 ، أظهر دبليو كانون أن قدرة الجسم على الحفاظ على التوازن هي نتيجة الأنظمة التنظيمية في الجسم. كما صاغ مصطلح الاستتباب. إن ثبات البيئة الداخلية للجسم (الدم ، اللمف ، سوائل الأنسجة ، السيتوبلازم) واستقرار الوظائف الفسيولوجية هو نتيجة آليات التماثل الساكن. في انتهاك للاستتباب ، مثل الخلوي ، هناك تنكس أو موت للخلايا. يتم تنظيم وتنسيق الخلايا والأنسجة والأعضاء والأشكال الأخرى من التوازن عن طريق التنظيم الخلطي والعصبي ، فضلاً عن مستوى التمثيل الغذائي.
معلمات الاستتباب ديناميكية وتتغير ضمن حدود معينة تحت تأثير العوامل البيئية (على سبيل المثال ، درجة الحموضة في الدم ، ومحتوى غازات الجهاز التنفسي والجلوكوز فيه ، وما إلى ذلك). هذا يرجع إلى حقيقة أن الأنظمة الحية لا توازن بين التأثيرات الخارجية فحسب ، بل تتصدى لها بفعالية. القدرة على الحفاظ على ثبات البيئة الداخلية مع التغيرات في البيئة الخارجية هي الخاصية الرئيسية التي تميز الكائنات الحية عن الطبيعة غير الحية. لذلك ، فهي مستقلة تمامًا عن البيئة الخارجية. كلما زاد تنظيم الكائن الحي ، زاد استقلاليته عن البيئة الخارجية (مثال).
يسمى مجمع العمليات التي توفر التوازن بالحركة المنزلية. يتم إجراؤه بواسطة جميع أنسجة وأعضاء وأنظمة الجسم. ومع ذلك ، فإن الأنظمة الوظيفية لها أهمية قصوى.
^ ملامح العمر للوظائف الفسيولوجية
والتنظيم العصبي
في عملية تطور الكائن الحي ، تحدث تغيرات كمية ونوعية. على سبيل المثال ، يزيد عدد الخلايا وأحجامها. في الوقت نفسه ، نتيجة لتعقيد بنية الجسم ، تظهر وظائف جديدة. على سبيل المثال ، يكتسب دماغ الطفل النامي القدرة على التفكير المجرد.
الظواهر التالية تكمن وراء التغيرات المرتبطة بالعمر في وظائف أجهزة الجسم:
1. النضج غير المتكافئ أو غير المتزامن لأعضاء وأنظمة الجسم. 2 . العمر المرحلي يقفز.
3. التسريع. أولئك. تسريع معدل التطور البيولوجي في فترة معينة.
لا يحدث نضج الأعضاء والأنظمة الفردية في وقت واحد (متغاير التوقيت). في حديثي الولادة ، أولاً وقبل كل شيء ، تتطور تلك الأنظمة الفسيولوجية والوظيفية التي تضمن بقاء الكائن الحي خلال هذه الفترة الانتقال من داخل الرحم إلى وجود خارج الرحم. بناءً على ملاحظات تكوين الأنظمة الوظيفية في عملية التولد ، ذكر الأكاديمي P.K. ابتكر أنوخين عقيدة تكوين النظام. يمكن توضيح التغاير الزمني لتطور الأعضاء والأنظمة من خلال مثال الجهاز الحركي للطفل. في البداية ، تتشكل ردود الفعل التي توفر إمساك الرأس ، ثم القدرة على الجلوس ، ثم الوقوف ، وأخيرًا المشي. برنامج التنمية الفرديةالتي يؤديها الجهاز الجيني. في مراحل عمرية معينة ، يحدث التعبير ، أي تنشيط الجينات المحددة جيدًا. نتيجة لذلك ، يتم تسريع نضج نظام معين أو وظيفة الجسم. يتجلى ذلك في قفزة عمرية أو فترة حرجة. على سبيل المثال ، لوحظ تغيير مفاجئ في بنية ووظيفة الأعضاء والأنظمة خلال فترة البلوغ.
يرتبط التسارع بتأثير العوامل البيئية والاجتماعية على الجسم. يترافق مع نمو سريع للهيكل العظمي والعضلات ، اعضاء داخلية، سن البلوغ.
ينتهي تكوين الجسم وتطوره بحوالي 20 عامًا. يعتبر الأشخاص الذين تتراوح أعمارهم بين 20 إلى 55-60 سنة من كبار السن ، وخلال هذه الفترة تتشكل جميع وظائف الجسم بشكل كامل ، نشاط وظيفيالأجهزة والأنظمة على نفس المستوى تقريبًا. بالنسبة لكبار السن الذين تتراوح أعمارهم بين 65 و 75 عامًا ، فإن حدوث إعادة الترتيب اللاإرادي هو سمة مميزة. واحدة من العلامات الرئيسية للشيخوخة هو انخفاض التمثيل الغذائي القاعدي ، ونتيجة لذلك تتعطل عمليات التمثيل الغذائي في الخلايا. ينخفض التمثيل الغذائي الأساسي نتيجة لانخفاض عدد الميتوكوندريا في الخلايا. يُعتقد أن قيمة التمثيل الغذائي القاعدي من أهم العوامل التي تحدد مدة حياة الإنسان. بعد 75 عامًا ، تبدأ الشيخوخة. نشاط جميع العمليات الفسيولوجية ينخفض بشكل حاد. نتيجة لذلك ، تحدث العديد من أمراض الشيخوخة ، مثل تصلب الشرايين.
تتغير آليات التنظيم العصبي العصابي أيضًا مع تقدم العمر. لدى المولود عدد محدود من ردود الفعل المعقدة غير المشروطة ، ولا توجد ردود فعل مشروطة. في الوقت نفسه ، تكون الخلايا شديدة الحساسية للعوامل الخلطية. مع النمو ، يتحسن النشاط الانعكاسي للجهاز العصبي المركزي. على وجه الخصوص ، بحلول عام واحد ، تتشكل ردود أفعال معقدة للغاية توفر الكلام. في الوقت نفسه ، تنخفض الحساسية الأولية للخلايا للعوامل الخلطية. لدى الشخص الناضج آليات عالية التنظيم للتنظيم العصبي. في الشيخوخة ، تقل سرعة وشدة ردود الفعل الانعكاسية. ضعف التأثيرات العصبية على الأعضاء والأنسجة ناتج عن التغيرات المدمرة في النهايات العصبية والمشابك العصبية في الجهاز العصبي المركزي والمحيط. في الوقت نفسه ، بسبب التغيرات في جهاز المستقبل للخلايا ، تقل قابليتها لعدد من العوامل الخلطية.
بالنسبة لأعضاء هيئة التدريس في طب الأطفال ، من المهم معرفة فترات الطفولة. تخصيص (حسب أرشافسكي):
1. فترة حديثي الولادة هي 7-8 أيام.
2. الفترة الرضاعة الطبيعية- 5-6 شهور
3. فترة التغذية المختلطة من 6 إلى 12 شهر.
4. فترة الطفل 1 سنة - 3 سنوات
5. فترة ما قبل المدرسة سن 3 - 7 سنوات.
6. فترة الدراسة الابتدائية سن 7-12 سنة
7. فترة المدرسة الثانوية سن 12-17 سنة
8 سنوات الشباب 17-20 سنة
PHYS I OLOGY AND B I O PHYS I C A V O Z B U D I M X
C L E T O C
^ مفهوم التهيج والاستثارة والإثارة. تصنيف المنبهات
التهيج هو قدرة الخلايا والأنسجة والجسم ككل على التحرك تحت تأثير العوامل البيئية الخارجية أو الداخلية من حالة الراحة الفسيولوجية إلى حالة النشاط. تتجلى حالة النشاط من خلال تغيير في المعلمات الفسيولوجية لخلية أو نسيج أو كائن حي ، على سبيل المثال ، تغيير في التمثيل الغذائي.
الاستثارة هي قدرة الأنسجة الحية على الاستجابة للتهيج برد فعل محدد نشط - الإثارة ، أي توليد النبض العصبي والتقلص والإفراز. أولئك. تتميز استثارة الأنسجة المتخصصة - العصبية والعضلية والغدية ، والتي تسمى استثارة. الإثارة عبارة عن مجموعة من عمليات استجابة النسيج المثير لتأثير مادة مهيجة ، ويتجلى ذلك من خلال تغيير في إمكانات الغشاء ، والتمثيل الغذائي ، وما إلى ذلك. الأنسجة المثيرة هي موصلة. هذه هي قدرة الأنسجة على إجراء الإثارة. تتمتع الأعصاب والعضلات الهيكلية بأعلى موصلية.
المهيج هو عامل البيئة الخارجية أو الداخلية التي تؤثر على الأنسجة الحية.
تسمى عملية تعرض مادة مهيجة لخلية أو نسيج أو كائن حي بالتهيج.
تنقسم جميع المحفزات إلى المجموعات التالية:
1. بحكم طبيعتها
أ) المادية (الكهرباء ، الضوء ، الصوت ، التأثيرات الميكانيكية ، إلخ)
ب) مادة كيميائية (أحماض ، قلويات ، هرمونات ، إلخ)
ج) الفيزيائية والكيميائية (الضغط الاسموزي ، الضغط الجزئي للغازات ، إلخ)
د) بيولوجي (طعام لحيوان ، فرد من الجنس الآخر)
د) اجتماعي (كلمة لشخص).
2 - حسب مكان التأثير:
أ) خارجي (خارجي)
ب) داخلي (داخلي)
3-بالقوة:
أ) عتبة فرعية (لا تسبب استجابة)
ب) العتبة (محفزات الحد الأدنى من القوة التي تحدث عندها الإثارة)
ج) العتبة الفائقة (القوة فوق العتبة)
4. بطبيعتها الفسيولوجية:
أ) مناسب (فسيولوجي لخلية أو مستقبلات معينة تكيفت معها في عملية التطور ، على سبيل المثال ، الضوء لمستقبلات العين الضوئية).
ب) غير كاف
إذا كان رد الفعل على المنبه منعكسًا ، فهناك أيضًا:
أ) المنبهات المنعكسة غير المشروطة
ب) المنعكس الشرطي
^ قوانين التهيج. معلمات استثارة.
يتم تحديد رد فعل الخلايا والأنسجة على مهيج من خلال قوانين التهيج
1. قانون "الكل أو لا شيء": مع تهيج الخلايا قبل العتبة ، لا تحدث استجابة الأنسجة. عند عتبة قوة التحفيز ، تتطور الاستجابة القصوى ، وبالتالي فإن زيادة قوة التهيج فوق العتبة لا تكون مصحوبة بزيادتها. وفقًا لهذا القانون ، يستجيب عصب وعضلة واحدة ، عضلة القلب ، للمنبهات.
2. 2. قانون القوة: كلما زادت قوة الحافز ، زادت الاستجابة. ومع ذلك ، فإن شدة الاستجابة تزيد فقط إلى حد أقصى معين. تخضع العضلات الهيكلية الشاملة والملساء لقانون القوة ، لأنها تتكون من خلايا عضلية عديدة ذات استثارة مختلفة.
3. قانون مدة القوة. هناك علاقة معينة بين قوة التحفيز ومدته. كلما كان الحافز أقوى ، قل الوقت الذي تستغرقه الاستجابة. تنعكس العلاقة بين قوة العتبة والمدة المطلوبة للتحفيز في منحنى مدة القوة. يمكن تحديد عدد من معلمات استثارة من هذا المنحنى.أ) عتبة التهيج هي الحد الأدنى من قوة التحفيز الذي يحدث عنده الإثارة.
ب) Reobase هو الحد الأدنى من قوة المنبه الذي يسبب الإثارة أثناء عمله لفترة طويلة إلى أجل غير مسمى. من الناحية العملية ، فإن العتبة وقاعدة الريباسي لهما نفس المعنى. وكلما انخفضت عتبة التهيج أو قلَّت التهابات ، زادت استثارة الأنسجة.
ج) الوقت المفيد - هذا هو الحد الأدنى من وقت عمل المنبه بقوة من ريوباز واحد يحدث خلاله الإثارة.
د) كروناكسيا - هذا هو الحد الأدنى من وقت عمل التحفيز بقوة من قاعدتين ريوبازيتين ، وهو أمر ضروري لبدء الإثارة. تم اقتراح هذه المعلمة ليتم حسابها بواسطة L. Lapik ، من أجل تحديد أكثر دقة لمؤشر الوقت على منحنى مدة القوة. أقصر وقت مفيدأو الكروناكسيا ، كلما زادت الإثارة والعكس صحيح.
في الممارسة السريرية ، يتم تحديد الريوباز والكرونكسيا باستخدام طريقة الكرونكسيس لدراسة استثارة جذوع الأعصاب.
4. قانون التدرج أو الإقامة. تعتمد استجابة الأنسجة للتهيج على تدرجها ، أي كلما زادت قوة التحفيز بمرور الوقت ، زادت سرعة الاستجابة. مع معدل منخفض من الزيادة في قوة التحفيز ، تزداد عتبة التهيج. لذلك ، إذا زادت قوة المنبه ببطء شديد ، فلن يكون هناك إثارة. هذه الظاهرة تسمى الإقامة.
القدرة الفسيولوجية (التنقل) هي تكرار أكبر أو أقل من ردود الفعل التي يمكن أن تستجيب الأنسجة للتحفيز الإيقاعي. كلما تمت استعادة استثارته بشكل أسرع بعد التهيج التالي ، زادت قابليته للتهيج. تم اقتراح تعريف lability من قبل N.E. ففيدنسكي. أعظم ضعف في الأعصاب ، وأصغر في عضلة القلب.
^ عمل التيار المباشر على الأنسجة القابلة للإثارة
لأول مرة ، تمت دراسة انتظام عمل التيار المباشر على عقار عصبي عضلي في القرن التاسع عشر بواسطة Pfluger. وجد أنه عند إغلاق دائرة التيار المستمر ، تحت القطب السالب ، أي تزيد الاستثارة تحت الكاثود وتنخفض تحت القطب الموجب. هذا يسمى قانون التيار المباشر. يسمى التغيير في استثارة الأنسجة (على سبيل المثال ، العصب) تحت تأثير تيار مباشر في منطقة الأنود أو الكاثود ، النغمة الكهربائية الفسيولوجية. لقد ثبت الآن أنه تحت تأثير القطب السالب - الكاثود ، تقل إمكانات غشاء الخلية. هذه الظاهرة تسمى catelectroton الفيزيائية. تحت الأنود الموجب ، يزداد. هناك anelektrton الجسدية. نظرًا لأن إمكانات الغشاء ، تحت الكاثود ، تقترب من المستوى الحرج لإزالة الاستقطاب ، تزداد استثارة الخلايا والأنسجة. تحت الأنود ، تزداد إمكانات الغشاء وتتحرك بعيدًا عن المستوى الحرج لإزالة الاستقطاب ، وبالتالي تقل استثارة الخلية والأنسجة. وتجدر الإشارة إلى أنه مع إجراء قصير المدى للغاية للتيار المباشر (1 مللي ثانية أو أقل) ، فإن MP ليس لديه الوقت للتغيير ، وبالتالي ، فإن استثارة الأنسجة الموجودة تحت الأقطاب الكهربائية لا تتغير أيضًا.
يستخدم التيار المباشر على نطاق واسع في العيادة للعلاج والتشخيص. على سبيل المثال ، يتم استخدامه للتحفيز الكهربائي للأعصاب والعضلات ، والعلاج الطبيعي: الرحلان الشاردي والجلفنة.
^ هيكل ووظائف الغشاء السيتوبلازمي للخلايا.
يتكون غشاء الخلية السيتوبلازمي من ثلاث طبقات: طبقة بروتينية خارجية وطبقة شحمية ثنائية الجزيء وطبقة بروتينية داخلية. سمك الغشاء 7.5-10 نانومتر. الطبقة ثنائية الجزيئية من الدهون هي مصفوفة الغشاء. تتفاعل جزيئات الدهون في كلتا الطبقتين مع جزيئات البروتين المغمورة فيهما. من 60 إلى 75٪ من دهون الغشاء عبارة عن شحميات فسفورية ، 15-30٪ كوليسترول. يتم تمثيل البروتينات بشكل رئيسي بواسطة البروتينات السكرية. هناك بروتينات متكاملة تخترق الغشاء بأكمله والبروتينات الطرفية الموجودة على السطح الخارجي أو الداخلي. تشكل البروتينات المتكاملة قنوات أيونية تضمن تبادل أيونات معينة بين السائل خارج الخلايا وداخلها. وهي أيضًا إنزيمات تقوم بنقل الأيونات المضاد للتدرج عبر الغشاء. البروتينات المحيطية هي مستقبلات كيميائية على السطح الخارجي للغشاء ، والتي يمكن أن تتفاعل مع مختلف PAS.
وظائف الغشاء:
يضمن سلامة الخلية كوحدة هيكلية للنسيج.
يقوم بتبادل الأيونات بين السيتوبلازم والسائل خارج الخلية.
4. ينتج إدراك ومعالجة المعلومات الواردة إلى الخلية في شكل إشارات كيميائية وكهربائية.
^ آليات استثارة الخلية. القنوات الأيونية الغشائية.
آليات حدوث إمكانات الغشاء (MP) وإمكانات العمل (AP)
في الأساس ، تكون المعلومات المنقولة في الجسم في شكل إشارات كهربائية (على سبيل المثال ، نبضات عصبية). ولأول مرة ، تم إثبات وجود الكهرباء الحيوانية من قبل عالم وظائف الأعضاء L. Galvani في عام 1786. من أجل دراسة كهرباء الغلاف الجوي ، قام بتعليق المستحضرات العصبية العضلية لأرجل الضفادع على خطاف نحاسي. عندما لامست هذه الكفوف الدرابزين الحديدي للشرفة ، تقلصت العضلات. يشير هذا إلى عمل نوع من الكهرباء على عصب التحضير العصبي العضلي. اعتبر جالفاني أن هذا يرجع إلى وجود الكهرباء في الأنسجة الحية نفسها. ومع ذلك ، وجد A. Volta أن مصدر الكهرباء هو مكان التلامس بين معدنين مختلفين - النحاس والحديد. في علم وظائف الأعضاء ، تعتبر أول تجربة كلاسيكية لجالفاني أنها تلامس العصب بتحضير عصبي عضلي بملاقط ثنائية المعدن مصنوعة من النحاس والحديد. لإثبات قضيته ، أجرى جالفاني تجربة ثانية. ألقى نهاية العصب الذي يعصب التحضير العصبي العضلي على قطع عضلاته. كانت النتيجة انكماش. ومع ذلك ، فإن هذه التجربة لم تقنع معاصري جالفاني. لذلك ، قام ماتيوتشي إيطالي آخر بإجراء التجربة التالية. قام بتركيب عصب تحضير ضفدع عصبي عضلي على عضلة الضفدع الثاني ، والتي تقلصت تحت تأثير تيار مزعج. نتيجة لذلك ، بدأ الدواء الأول أيضًا في الانخفاض. يشير هذا إلى انتقال الكهرباء (PD) من عضلة إلى أخرى. تم تحديد وجود فرق الجهد بين الأجزاء التالفة وغير التالفة من العضلات بدقة لأول مرة في القرن التاسع عشر باستخدام مقياس الجلفانومتر الخيطي (مقياس التيار) Matteuchi. علاوة على ذلك ، كان للشق شحنة سالبة ، وكان سطح العضلة موجبًا.
^ تصنيف وهيكل القنوات الأيونية في السيتوبلازم
أغشية. آليات حدوث الغشاء المحتمل
وإمكانات العمل.
تم اتخاذ الخطوة الأولى في دراسة أسباب استثارة الخلية في عمله The Theory of Membrane Equilibrium في عام 1924 من قبل عالم وظائف الأعضاء الإنجليزي دونان. لقد أثبت نظريًا أن فرق الجهد داخل الخلية وخارجها ، أي الراحة المحتملة أو MP ، قريبة من إمكانات توازن البوتاسيوم. هذه هي الإمكانات المتكونة على محاليل فصل غشاء شبه منفذة بتركيزات مختلفة من أيونات البوتاسيوم ، يحتوي أحدها على أنيونات كبيرة غير قابلة للاختراق. حدد نيرنست حساباته. اشتق معادلة الانتشار المحتملة. بالنسبة للبوتاسيوم ، سيكون مساويًا لـ:
Ек = 58 lg -------- = 58 lg ----- = - 75 مللي فولت ،
هذه هي القيمة المحسوبة نظريًا لـ MP.
تجريبيًا ، تم إنشاء آليات ظهور فرق محتمل بين السائل خارج الخلية والسيتوبلازم ، وكذلك إثارة الخلايا ، في عام 1939 في كامبريدج بواسطة هودجكين وهكسلي. قاموا بفحص الألياف العصبية العملاقة (محور عصبي) للحبار ووجدوا أن السائل داخل الخلايا للخلايا العصبية يحتوي على 400 ملي مولار من البوتاسيوم ، و 50 ملي مولار من الصوديوم ، و 100 ملي كلور ، وقليل جدًا من الكالسيوم. احتوى السائل خارج الخلية على 10 ملي مولار من البوتاسيوم ، و 440 ملي مولار من الصوديوم ، و 560 ملي مولار من الكلور ، و 10 ملي مولار من الكالسيوم. وبالتالي ، يوجد فائض من البوتاسيوم داخل الخلايا والصوديوم والكالسيوم خارجها. ويرجع ذلك إلى حقيقة أن القنوات الأيونية مدمجة في غشاء الخلية ، والتي تنظم نفاذية غشاء أيونات الصوديوم والبوتاسيوم والكالسيوم والكلور.
جميع القنوات الأيونية مقسمة إلى المجموعات التالية:
1. بالانتقائية:
أ) انتقائي ، أي محدد. هذه القنوات قابلة للاختراق للأيونات المحددة بدقة.
ب) انتقائية منخفضة ، غير محددة ، ليس لها انتقائية أيون معينة. لا يوجد سوى عدد قليل منهم في الغشاء.
2. حسب طبيعة الأيونات المرسلة:
أ) البوتاسيوم
ب) الصوديوم
ب) الكالسيوم
د) الكلور
3. حسب معدل التعطيل ، أي إغلاق:
أ) التعطيل السريع ، أي تتحول بسرعة إلى حالة مغلقة. أنها توفر انخفاضًا سريعًا متزايدًا في MP ونفس الانتعاش السريع.
ب) التعطيل ببطء. يؤدي افتتاحها إلى انخفاض بطيء في MP وتعافيها البطيء.
آليات الفتح:
ب) تعتمد على المواد الكيميائية ، وتفتح عندما تتعرض المستقبلات الكيميائية لغشاء الخلية للمواد الفعالة فيزيولوجيًا (النواقل العصبية والهرمونات وما إلى ذلك).
لقد ثبت الآن أن القنوات الأيونية لها الهيكل التالي:
1. مرشح انتقائي يقع عند فم القناة. يضمن مرور الأيونات المحددة بدقة عبر القناة.
2. بوابات التنشيط التي تفتح عند مستوى معين من إمكانات الغشاء أو عمل PAS المقابل. تحتوي بوابات تنشيط القنوات ذات الجهد الكهربائي على مستشعر يفتحها عند مستوى MP معين.
3. بوابة التعطيل ، والتي تضمن إغلاق القناة وإنهاء توصيل الأيونات عبر القناة عند مستوى معين من MP (الشكل).
لا تحتوي القنوات الأيونية غير المحددة على بوابة.
يمكن أن تكون القنوات الأيونية الانتقائية في ثلاث حالات ، والتي يتم تحديدها من خلال موضع التنشيط (م) وتعطيل (ح) بوابات (الشكل):
1. يتم إغلاقها عند إغلاق ملفات التنشيط وفتح ملفات التنشيط.
2-مفعلة كلا البوابتين مفتوحتان.
3. تعطيل بوابات التفعيل وإغلاق بوابات التعطيل.
يتم تحديد الموصلية الكلية لأيون معين من خلال عدد القنوات المقابلة المفتوحة في نفس الوقت. في حالة الراحة ، يتم فتح قنوات البوتاسيوم فقط ، مما يحافظ على إمكانات غشاء معينة ، ويتم إغلاق قنوات الصوديوم. لذلك ، يكون الغشاء نافذًا انتقائيًا للبوتاسيوم وقليلًا جدًا لأيونات الصوديوم والكالسيوم ، بسبب وجود قنوات غير محددة. نسبة نفاذية الغشاء للبوتاسيوم والصوديوم في حالة الراحة هي 1: 0.04. تدخل أيونات البوتاسيوم إلى السيتوبلازم وتتراكم فيه. عندما يصل عددهم إلى حد معين ، يبدأون في مغادرة الخلية عبر قنوات البوتاسيوم المفتوحة على طول تدرج التركيز. ومع ذلك ، لا يمكنهم الهروب من السطح الخارجي لغشاء الخلية. هناك يتم احتجازهم بواسطة المجال الكهربائي للأنيونات سالبة الشحنة الموجودة على السطح الداخلي. هذه هي أنيونات الكبريتات والفوسفات والنترات ، وهي مجموعات أنيونية من الأحماض الأمينية التي يكون الغشاء غير منفذ لها. لذلك ، تتراكم كاتيونات البوتاسيوم موجبة الشحنة على السطح الخارجي للغشاء ، وتتراكم الأنيونات سالبة الشحنة على السطح الداخلي. هناك فرق جهد عبر الغشاء. أرز.
يحدث إطلاق أيونات البوتاسيوم من الخلية حتى يوازن الجهد الناشئ مع علامة موجبة من الخارج تدرج تركيز البوتاسيوم الموجه خارج الخلية. أولئك. أيونات البوتاسيوم المتراكمة على الجانب الخارجي من الغشاء لن تطرد نفس الأيونات إلى الداخل. تنشأ إمكانات غشاء معينة ، يتم تحديد مستواها بواسطة موصلية الغشاء لأيونات البوتاسيوم والصوديوم أثناء الراحة. في المتوسط ، تكون قيمة إمكانات الراحة قريبة من إمكانات توازن البوتاسيوم Nernst. على سبيل المثال ، MP للخلايا العصبية هو 55-70 mV ، مخطط - 90-100 mV ، العضلات الملساء - 40-60 mV ، الخلايا الغدية - 20-45 mV. تفسر القيمة الحقيقية المنخفضة لـ MP للخلايا من خلال حقيقة أن قيمتها تقل عن طريق أيونات الصوديوم ، والتي يكون الغشاء فيها منفذاً قليلاً ويمكنها دخول السيتوبلازم. من ناحية أخرى ، تؤدي أيونات الكلوريد السالبة التي تدخل الخلية إلى زيادة طفيفة في MP.
نظرًا لأن الغشاء السكون يكون منفذاً قليلاً لأيونات الصوديوم ، فهناك حاجة إلى آلية لإزالة هذه الأيونات من الخلية. هذا يرجع إلى حقيقة أن التراكم التدريجي للصوديوم في الخلية من شأنه أن يؤدي إلى تحييد إمكانات الغشاء واختفاء الاستثارة. هذه الآلية تسمى مضخة الصوديوم والبوتاسيوم. يحافظ على الفرق بين تركيزات البوتاسيوم والصوديوم على جانبي الغشاء. مضخة الصوديوم والبوتاسيوم هي إنزيم الصوديوم والبوتاسيوم ATPase. يتم تضمين جزيئات البروتين في الغشاء. يقوم بتكسير ATP ويستخدم الطاقة المنبعثة لإزالة الصوديوم بالتدرج المعاكس من الخلية وضخ البوتاسيوم فيها. في دورة واحدة ، يزيل كل جزيء ATPase الصوديوم والبوتاسيوم 3 أيونات الصوديوم ويساهم في 2 أيونات البوتاسيوم. نظرًا لأن عدد الأيونات الموجبة الشحنة تدخل الخلية أقل مما يتم إزالته منها ، فإن الصوديوم والبوتاسيوم ATPase يزيد من إمكانات الغشاء بمقدار 5-10 مللي فولت.
يحتوي الغشاء على الآليات التالية لنقل الأيونات والمواد الأخرى عبر الغشاء:
1. النقل النشط. يتم تنفيذه باستخدام طاقة ATP. تشمل هذه المجموعة من أنظمة النقل مضخة الصوديوم والبوتاسيوم ومضخة الكالسيوم ومضخة الكلور.
2. النقل السلبي. تتم حركة الأيونات على طول تدرج التركيز دون استهلاك الطاقة. على سبيل المثال دخول البوتاسيوم إلى الخلية والخروج منها عبر قنوات البوتاسيوم.
3. النقل المرتبط. النقل المضاد للتدرج للأيونات بدون استهلاك للطاقة. على سبيل المثال ، يحدث التبادل الأيوني للصوديوم والصوديوم والصوديوم والكالسيوم والبوتاسيوم والبوتاسيوم بهذه الطريقة. يحدث بسبب الاختلاف في تركيز الأيونات الأخرى.
يتم تسجيل جهد الغشاء باستخدام طريقة المسرى الدقيق. للقيام بذلك ، يتم إدخال قطب كهربائي دقيق رفيع ، أقل من 1 ميكرومتر من الزجاج من خلال الغشاء في سيتوبلازم الخلية. إنها مليئة بمحلول ملحي. يتم وضع القطب الثاني في السائل المحيط بالخلايا. من الأقطاب الكهربائية ، تنتقل الإشارة إلى مضخم الجهد الحيوي ، ومنه إلى الذبذبات والمسجل (الشكل).
أظهرت دراسات أخرى أجراها هودجكين وهكسلي أنه عندما يكون محور الحبار متحمسًا ، يحدث تقلب سريع في إمكانات الغشاء ، والذي كان له شكل ذروة (ارتفاع) على شاشة راسم الذبذبات. أطلقوا على هذا التذبذب إمكانات الفعل (AP). نظرًا لأن التيار الكهربائي للأغشية القابلة للاستثارة هو حافز مناسب ، يمكن تحفيز AP عن طريق وضع قطب كهربائي سالب ، الكاثود ، على السطح الخارجي للغشاء ، وأنود على السطح الداخلي الموجب. سيؤدي هذا إلى انخفاض في قيمة شحنة الغشاء - إزالة الاستقطاب. تحت تأثير تيار عتبة فرعية ضعيف ، يحدث نزع الاستقطاب السلبي ، أي يحدث catelectroton (الشكل). إذا زادت القوة الحالية إلى حد معين ، فعند نهاية فترة تأثيرها على هضبة catelectroton ، سيظهر ارتفاع تلقائي صغير - استجابة محلية أو محلية. إنه نتيجة لفتح جزء صغير من قنوات الصوديوم الموجودة أسفل الكاثود. مع تيار عتبة ، ينخفض MP إلى المستوى الحرج لإزالة الاستقطاب (CDL) ، حيث يبدأ توليد جهد فعل. إنه تقريبًا عند مستوى -50 مللي فولت للخلايا العصبية.
يتم تمييز المراحل التالية على منحنى جهد الفعل:
1. الاستجابة المحلية (نزع الاستقطاب المحلي) التي سبقت تطور شلل الرعاش.
2. مرحلة نزع الاستقطاب. خلال هذه المرحلة ، يتناقص MF بسرعة ويصل إلى الصفر. يرتفع مستوى إزالة الاستقطاب إلى ما فوق الصفر. لذلك ، يكتسب الغشاء شحنة معاكسة - داخله تصبح موجبة ، وخارجه تصبح سالبة. تسمى ظاهرة تغيير شحنة الغشاء ارتداد جهد الغشاء. مدة هذه المرحلة في الخلايا العصبية والعضلية هي 1-2 مللي ثانية.
3. مرحلة عودة الاستقطاب. يبدأ عند الوصول إلى مستوى معين من MP (حوالي +20 مللي فولت). تبدأ إمكانات الغشاء في العودة بسرعة إلى إمكانات الراحة. مدة المرحلة 3-5 مللي ثانية.
4. مرحلة إزالة الاستقطاب التتبع أو تتبع الإمكانات السلبية. الفترة التي يتم فيها تأخير عودة النائب إلى إمكانات الراحة مؤقتًا. يستمر 15-30 مللي ثانية.
5. مرحلة تتبع فرط الاستقطاب أو تتبع إمكانات إيجابية. في هذه المرحلة ، يصبح MP لبعض الوقت أعلى من المستوى الأولي لـ PP. مدته 250-300 مللي ثانية.
تبلغ سعة إمكانات عمل العضلات الهيكلية في المتوسط 120-130 مللي فولت ، والخلايا العصبية 80-90 مللي فولت ، وخلايا العضلات الملساء 40-50 مللي فولت. عندما تكون الخلايا العصبية متحمسة ، تحدث AP في الجزء الأولي من المحور العصبي - التل المحوري.
يرجع حدوث AP إلى تغيير في النفاذية الأيونية للغشاء عند الإثارة. خلال فترة الاستجابة المحلية ، تفتح قنوات الصوديوم البطيئة ، بينما تبقى القنوات السريعة مغلقة ، ويحدث نزع الاستقطاب التلقائي المؤقت. عندما يصل MP إلى مستوى حرج ، تفتح بوابات التنشيط المغلقة لقنوات الصوديوم وتندفع أيونات الصوديوم إلى الخلية مثل الانهيار الجليدي ، مما يتسبب في إزالة الاستقطاب التدريجي. يتم فتح قنوات الصوديوم السريعة والبطيئة خلال هذه المرحلة. أولئك. تزداد نفاذية الصوديوم في الغشاء بشكل حاد. علاوة على ذلك ، فإن قيمة المستوى الحرج لإزالة الاستقطاب تعتمد على حساسية التنشيط ، فكلما كانت أعلى ، انخفض FCA والعكس صحيح.
عندما تقترب كمية إزالة الاستقطاب من احتمال توازن أيونات الصوديوم (+20 mV). يتم تقليل قوة انحدار تركيز الصوديوم بشكل كبير. في الوقت نفسه ، تبدأ عملية تعطيل قنوات الصوديوم السريعة وانخفاض موصلية الصوديوم للغشاء. توقف الاستقطاب. يزداد إنتاج أيونات البوتاسيوم بشكل حاد ، أي تيار انتاج البوتاسيوم. في بعض الخلايا ، يرجع ذلك إلى تنشيط قنوات تدفق البوتاسيوم الخاصة. يعمل هذا التيار ، الموجه من الخلية ، على نقل MP بسرعة إلى مستوى إمكانات الراحة. أولئك. تبدأ مرحلة عودة الاستقطاب. تؤدي الزيادة في MP إلى إغلاق بوابات تنشيط قنوات الصوديوم ، مما يقلل من نفاذية الصوديوم في الغشاء ويسرع عودة الاستقطاب.
يمكن تفسير حدوث مرحلة إزالة الاستقطاب من التتبع بحقيقة عدم حدوث ذلك معظمتبقى قنوات الصوديوم البطيئة مفتوحة.
يرتبط تتبع فرط الاستقطاب بزيادة توصيل البوتاسيوم للغشاء بعد PD ، وحقيقة أن مضخة الصوديوم والبوتاسيوم أكثر نشاطًا ، حيث تقوم بتنفيذ أيونات الصوديوم التي دخلت الخلية أثناء PD.
من خلال تغيير موصلية قنوات الصوديوم والبوتاسيوم السريعة ، من الممكن التأثير على توليد AP ، وبالتالي إثارة الخلايا. مع الحصار الكامل لقنوات الصوديوم ، على سبيل المثال ، مع سم سمكة tetrodont - tetrodotoxin ، تصبح الخلية غير مهتمة. يستخدم هذا في العيادة. مثل تخدير موضعي، كما نوفوكائين ، ديكايين ، ليدوكائين يمنع انتقال قنوات الصوديوم الألياف العصبيةللدولة المفتوحة. لذلك ، يتوقف توصيل النبضات العصبية على طول الأعصاب الحسية ، ويحدث تخدير (تخدير) للعضو. مع الحصار المفروض على قنوات البوتاسيوم ، يصعب إطلاق أيونات البوتاسيوم من السيتوبلازم إلى السطح الخارجي للغشاء ؛ انتعاش MP. لذلك ، يتم إطالة مرحلة عودة الاستقطاب. يستخدم هذا التأثير لحاصرات قنوات البوتاسيوم أيضًا في الممارسة السريرية. على سبيل المثال ، أحدها ، الكينيدين ، يطيل مرحلة عودة استقطاب خلايا عضلة القلب ، ويبطئ تقلصات القلب ويعيد إيقاع القلب إلى طبيعته.
وتجدر الإشارة أيضًا إلى أنه كلما زاد معدل انتشار AP على طول غشاء الخلية أو الأنسجة ، زادت موصليةها.
^ نسبة أطوار إمكانية العمل والإثارة
يعتمد مستوى استثارة الخلية على مرحلة AP. في مرحلة الاستجابة المحلية ، تزيد الإثارة. تسمى هذه المرحلة من استثارة الإضافة الكامنة.
في مرحلة إعادة الاستقطاب AP ، عندما تنفتح جميع قنوات الصوديوم وتندفع أيونات الصوديوم إلى الخلية مثل الانهيار الجليدي ، لا يمكن حتى لأي محفز فائق القوة أن يحفز هذه العملية. لذلك ، فإن مرحلة إزالة الاستقطاب تتوافق مع مرحلة عدم الاستثارة الكاملة أو الانكسار المطلق.
خلال مرحلة عودة الاستقطاب ، يتم إغلاق المزيد والمزيد من قنوات الصوديوم. ومع ذلك ، يمكن إعادة فتحها تحت تأثير حافز فوقي. أولئك. تبدأ الإثارة في الارتفاع مرة أخرى. هذا يتوافق مع مرحلة عدم الاستثارة النسبية أو الحران النسبي.
أثناء إزالة الاستقطاب من التتبع ، يكون MP في مستوى حرج ، لذلك حتى منبهات ما قبل العتبة يمكن أن تسبب إثارة الخلية. لذلك ، في هذه اللحظة ، تزداد استثارتها. تسمى هذه المرحلة بمرحلة التمجيد أو استثارة فائقة.
في لحظة تتبع فرط الاستقطاب ، يكون MP أعلى من المستوى الأولي ، أي مزيد من KUD ويتم تقليل استثارته. إنها في مرحلة استثارة غير طبيعية. أرز. تجدر الإشارة إلى أن ظاهرة الإقامة ترتبط أيضًا بتغيير في توصيل القنوات الأيونية. إذا زاد تيار إزالة الاستقطاب ببطء ، فإن هذا يؤدي إلى تعطيل جزئي للصوديوم وتنشيط قنوات البوتاسيوم. لذلك ، لا يحدث تطور PD.
^ فيزيولوجيا العضلات
يوجد 3 أنواع من العضلات في الجسم: عضلية هيكلية أو مخططة ، ملساء وعضلات قلبية. توفر عضلات الهيكل العظمي حركة الجسم في الفضاء ، مما يحافظ على وضعية الجسم بسبب تناغم عضلات الأطراف والجسم. العضلات الملساء ضرورية لتمعج الجهاز الهضمي ، الجهاز البولي، تنظيم نبرة الأوعية الدموية ، القصبات الهوائية ، إلخ. تُستخدم عضلة القلب لتقلص القلب وضخ الدم. تتمتع جميع العضلات بالاستثارة والتوصيل والانقباض ، ولدى القلب والعديد من العضلات الملساء تلقائية - القدرة على الانقباضات التلقائية.
^ البنية التحتية للألياف العضلية الهيكلية.
الوحدات الحركية العنصر المورفو الوظيفي الرئيسي للجهاز العصبي العضلي للعضلات الهيكلية هو الوحدة الحركية. ويشمل العصبون الحركي للنخاع الشوكي مع ألياف عضلية يغذيها محوره. داخل العضلة ، يشكل هذا المحور عدة فروع طرفية. يشكل كل فرع من هذه الفروع جهة اتصال - مشابك عصبية عضلية على ألياف عضلية منفصلة. تسبب النبضات العصبية القادمة من الخلايا العصبية الحركية تقلصات لمجموعة معينة من ألياف العضلات.
تتكون عضلات الهيكل العظمي من حزم عضلية تتكون من عدد كبير من ألياف العضلات. كل ليف عبارة عن خلية أسطوانية يبلغ قطرها 10-100 ميكرون وطولها من 5 إلى 400 ميكرون. لها غشاء خلوي - غمد الليف العضلي. يحتوي الساركوبلازم على العديد من النوى والميتوكوندريا وتشكيلات الشبكة الساركوبلازمية (SR) وعناصر مقلصة - اللييفات العضلية. تحتوي الشبكة الساركوبلازمية على بنية غريبة. يتكون من نظام من الأنابيب المستعرضة والطولية والخزانات. الأنابيب المستعرضة هي امتداد للساركوبلازم في الخلية. تجاورهم أنابيب طولية مع صهاريج. نتيجة لهذا ، يمكن أن ينتشر جهد الفعل من غمد الليف العضلي إلى نظام شبكية الهيولى العضلية. تحتوي الألياف العضلية على أكثر من 1000 ليفية عضلية موجودة على طولها. يتكون كل ليف عضلي من 2500 ليفية أو خيوط عضلية. هذه خيوط من البروتينات الانقباضية الأكتين والميوسين. اللييفات الأولية من الميوسين سميكة ، ولييفات الأكتين الأولية رقيقة.
توجد على خيوط الميوسين عمليات عرضية برؤوس ممتدة بزاوية. في ألياف العضلات والهيكل العظمي ، يُظهر الفحص المجهري الضوئي وجود خط عرضي ، أي بالتناوب بين الخطوط الفاتحة والداكنة. تسمى العصابات المظلمة الأقراص A أو متباينة الخواص ، والأقراص I الخفيفة (الخواص). تتركز خيوط الميوسين في الأقراص A ، والتي لها تباين وبالتالي لها لون غامق. تتكون الأقراص I من خيوط أكتين. تظهر لوحة Z رفيعة في وسط الأقراص على شكل حرف "I". يتم إرفاق الألياف الأولية الأكتين به. يسمى قسم اللييف العضلي بين صفائحتين Z بالقسيم العضلي. إنه عنصر هيكلي في اللييفات العضلية. في حالة الراحة ، تدخل خيوط الميوسين السميكة الفجوات بين خيوط الأكتين لمسافة قصيرة فقط. لذلك ، يوجد في الجزء الأوسط من القرص A منطقة H أخف ، حيث لا توجد خيوط أكتين. يُظهر الفحص المجهري الإلكتروني خطًا M رفيعًا جدًا في وسطه. يتكون من سلاسل من البروتينات الداعمة التي ترتبط بها اللييفات الأولية للميوسين (الشكل.
^ آليات تقلص العضلات
مع الفحص المجهري الضوئي ، لوحظ أنه في لحظة الانكماش ، لا ينخفض عرض القرص A ، ولكن تضيق أقراص I ومناطق H من الساركوميرات. باستخدام المجهر الإلكتروني ، وجد أن طول خيوط الأكتين والميوسين لا يتغير في وقت الانكماش. لذلك طور هكسلي وهانسون نظرية انزلاق الخيط. وفقًا لذلك ، تقصر العضلة نتيجة حركة خيوط الأكتين الرقيقة في الفجوات بين خيوط الميوسين. هذا يؤدي إلى تقصير كل قسيم عضلي يتكون من اللييفات العضلية. يرجع انزلاق الشعيرات إلى حقيقة أنه عند الانتقال إلى الحالة النشطة ، ترتبط رؤوس عمليات الميوسين بمراكز خيوط الأكتين وتتسبب في تحركها بالنسبة إلى نفسها (حركات السكتة الدماغية). لكن هذه هي المرحلة الأخيرة من آلية الانقباض بأكملها. يبدأ الانكماش بحقيقة أن PD يحدث في منطقة الصفيحة الطرفية للعصب الحركي. ينتشر بسرعة عالية على طول غمد الليف العضلي ويمر منه على طول نظام الأنابيب المستعرضة SR إلى الأنابيب الطولية والصهاريج. هناك إزالة الاستقطاب من غشاء الخزانات ويتم إطلاق أيونات الكالسيوم منها في الساركوبلازم. تحتوي خيوط الأكتين على جزيئات من بروتينين آخرين - تروبونين وتروبوميوسين. بتركيز منخفض (أقل من 10-8 م) من الكالسيوم ، أي في حالة الراحة ، يمنع تروبوميوسين ربط جسور الميوسين بخيوط الأكتين. عندما تبدأ أيونات الكالسيوم في ترك SR ، يغير جزيء التروبونين شكله بحيث يحرر مراكز الأكتين النشطة من التروبوميوسين. ترتبط رؤوس الميوسين بهذه المراكز ويبدأ الانزلاق بسبب الارتباط الإيقاعي وفصل الجسور المستعرضة بخيوط الأكتين. في هذه الحالة ، تتحرك الرؤوس بشكل إيقاعي على طول خيوط الأكتين إلى أغشية Z. تستغرق 50 دورة من هذا القبيل لتقلص العضلات بالكامل. يسمى انتقال الإشارة من الغشاء المثير إلى اللييفات العضلية بالاقتران الكهروميكانيكي. عندما يتوقف توليد AP وتعود إمكانات الغشاء إلى مستواها الأصلي ، تبدأ مضخة Ca (إنزيم Ca-ATPase) في العمل. يتم ضخ أيونات الكالسيوم مرة أخرى في صهاريج الشبكة الساركوبلازمية وينخفض تركيزها إلى أقل من 10-8 م. تكتسب جزيئات التروبونين شكلها الأصلي ويبدأ التروبوميوسين مرة أخرى في منع المراكز النشطة للأكتين. تنفصل رؤوس الميوسين عنها وتعود العضلات إلى حالتها الأصلية المهدئة بسبب المرونة.
^ طاقة تقلص العضلات
ATP هو مصدر الطاقة للتقلص والاسترخاء. تحتوي رؤوس الميوسين على مواقع تحفيزية تعمل على تكسير ATP إلى ADP والفوسفات غير العضوي. أولئك. الميوسين هو أيضًا إنزيم ATPase. يزداد نشاط الميوسين باعتباره ATPase بشكل ملحوظ عندما يتفاعل مع الأكتين. مع كل دورة من التفاعل بين الأكتين والرأس ، يقسم الميوسين جزيء ATP واحد. لذلك ، كلما زاد عدد الجسور في حالة نشطة ، كلما زاد انقسام ATP ، كان الانكماش أقوى. لتحفيز نشاط ATPase للميوسين ، يلزم وجود أيونات الكالسيوم ، والتي يتم إطلاقها من SR ، والتي تعزز إطلاق مراكز الأكتين النشطة من تروبوميوسين. ومع ذلك ، فإن توفير ATP في الخلية محدود. لذلك ، لتجديد احتياطيات ATP ، يتم استعادتها - إعادة التركيب. يتم إجراؤه لاهوائيًا وهوائيًا. يتم تنفيذ عملية إعادة التركيب اللاهوائي بواسطة أنظمة الفوسفاجين والجليكولتيك. يستخدم الأول احتياطيات فوسفات الكرياتين لاستعادة ATP. يتم تقسيمها إلى كرياتين وفوسفات ، والتي يتم نقلها إلى ADP بمساعدة الإنزيمات (ADP + P = ATP). يوفر نظام إعادة التركيب الفوسفاجي أكبر قوة تقلص ، ولكن بسبب الكمية الصغيرة من فوسفات الكرياتين في الخلية ، يعمل فقط لمدة 5-6 ثوان من الانكماش. يستخدم نظام تحلل السكر التكسير اللاهوائي للجلوكوز (الجليكوجين) إلى حمض اللاكتيك لإعادة تخليق ATP. يوفر كل جزيء جلوكوز استرداد ثلاثة جزيئات ATP. إن إمكانات الطاقة لهذا النظام أعلى من طاقة الفوسفاجين ، ولكن حتى يمكن أن تعمل كمصدر لطاقة الانكماش لمدة 0.5 - 2 دقيقة فقط. في الوقت نفسه ، يصاحب عمل نظام تحلل السكر تراكم حمض اللاكتيك في العضلات وانخفاض محتوى الأكسجين. مع العمل المطول ، مع زيادة الدورة الدموية ، تبدأ عملية إعادة تخليق ATP بمساعدة الفسفرة المؤكسدة ، أي هوائي. إن إمكانات الطاقة لنظام الأكسدة أكبر بكثير من غيرها. تحدث العملية بسبب أكسدة الكربوهيدرات والدهون. مع العمل المكثف ، تتأكسد الكربوهيدرات بشكل أساسي ، مع عمل معتدل ، الدهون. يتطلب الاسترخاء أيضًا طاقة ATP. بعد الموت ، يتناقص محتوى ATP في الخلايا بسرعة ، وعندما يصبح أقل من المستوى الحرج ، لا يمكن فصل جسور الميوسين المتقاطعة عن خيوط الأكتين (حتى التحلل الذاتي الإنزيمي لهذه البروتينات). صرامة الموت يحدث. ATP ضروري للاسترخاء لأنه يحافظ على عمل مضخة Ca.
^ الميكانيكا الحيوية لتقلصات العضلات.
الانكماش الفردي ، الجمع ، الكزاز.
عندما يتم تطبيق تهيج عتبة واحدة أو عتبة فوقية على العصب أو العضلات الحركية ، يحدث انقباض واحد. من خلال التسجيل الرسومي ، يمكن تمييز ثلاث فترات متتالية على المنحنى الناتج:
1. الفترة الكامنة. هذا هو الوقت من لحظة تطبيق التهيج إلى بداية الانكماش. مدته حوالي 1-2 مللي ثانية. خلال الفترة الكامنة ، يتم إنشاء AP ونشره ، ويتم تحرير الكالسيوم من SR ، ويتفاعل الأكتين مع الميوسين ، وما إلى ذلك.
2. فترة التقصير. اعتمادًا على نوع العضلات (سريعة أو بطيئة) ، تتراوح مدتها من 10 إلى 100 مللي ثانية.،
3- فترة الاسترخاء. مدته أطول إلى حد ما من تقصير. أرز.
في حالة الانقباض الفردي ، تكون العضلة قادرة على العمل لفترة طويلة دون إرهاق ، لكن قوتها ضئيلة. لذلك ، فإن مثل هذه الانقباضات نادرة في الجسم ، على سبيل المثال ، يمكن أن تنقبض العضلات الحركية السريعة بهذه الطريقة. في كثير من الأحيان ، يتم تلخيص الانقباضات المفردة.
الجمع هو إضافة تقلصتين متتاليتين عند تطبيق محفزات عتبة أو حد فوقي عليه ، ويكون الفاصل الزمني بينهما أقل من مدة الانكماش الفردي ، ولكنه يزيد عن مدة فترة المقاومة. هناك نوعان من التلخيص: الجمع الكامل وغير الكامل. يحدث الجمع غير الكامل إذا تم تطبيق التحفيز المتكرر على العضلات عندما بدأت بالفعل في الاسترخاء. يحدث التام عندما يؤثر التهيج المتكرر على العضلات قبل بدء فترة الاسترخاء ، أي في نهاية فترة التقصير (الشكل 1.2). اتساع الانكماش مع الجمع الكامل أعلى منه مع الجمع غير الكامل. إذا انخفض الفاصل الزمني بين تهيجين. على سبيل المثال ، ضع الثانية في منتصف فترة التقصير ، فلن يكون هناك جمع ، لأن العضلات في حالة مقاومة للحرارة.
كُزاز- هو تقلص طويل للعضلة ناتج عن تجميع عدة انقباضات فردية تتطور عند تطبيق سلسلة من المحفزات المتتالية عليها. يوجد نوعان من الكزاز: مسنن وناعم. يُلاحظ الكزاز المسنن إذا كان كل تهيج لاحق يؤثر على العضلات عندما بدأت بالفعل في الاسترخاء. أولئك. لوحظ جمع غير كامل (الشكل). يحدث الكزاز الأملس عندما يتم تطبيق كل منبه لاحق في نهاية فترة التقصير. أولئك. هناك مجموعة كاملة من الانقباضات الفردية و (الشكل). اتساع الكزاز الأملس أكبر من الكزاز المسنن. عادة ، تنقبض عضلات الإنسان في وضع الكزاز الأملس. تحدث خشنة مع علم الأمراض ، مثل رعاش اليد مع تسمم الكحول ومرض باركنسون.
^ تأثير التردد وقوة التنبيه على اتساع الانكماش
إذا قمت بزيادة وتيرة التهيج تدريجيًا ، فإن اتساع تقلص الكزاز يزداد. عند تردد معين ، سيصبح الحد الأقصى. هذا التردد يسمى الأمثل. زيادة أخرى في وتيرة التهيج مصحوبة بانخفاض في القوة.
انكماش كزازي. يسمى التردد الذي تبدأ عنده سعة الانقباض في الانخفاض بالتردد pessimal. عند التردد العالي جدًا من التحفيز ، لا تنقبض العضلات (الشكل). تم اقتراح مفهوم الترددات المثلى والمتشائم من قبل N.E. Vvedensky. وجد أن كل تحفيز لقوة عتبة أو قوة عابرة ، تسبب تقلصًا ، يغير في نفس الوقت استثارة العضلات. لذلك ، مع الزيادة التدريجية في وتيرة التنبيه ، يتم تحويل عمل النبضات بشكل متزايد إلى بداية فترة الاسترخاء ، أي مرحلة التمجيد. عند التردد الأمثل ، تعمل جميع النبضات على العضلات في مرحلة التمجيد ، أي زيادة الإثارة. لذلك ، فإن سعة التيتانوس هي الحد الأقصى. مع زيادة تواتر التنبيه ، يؤثر عدد متزايد من النبضات على العضلات في المرحلة المقاومة للحرارة. تقل سعة التيتانوس.
تتفاعل الألياف العضلية المفردة ، مثل أي خلية قابلة للاستثارة ، مع التهيج وفقًا لقانون الكل أو لا شيء. العضلة تخضع لقانون القوة. مع زيادة قوة التحفيز ، يزداد اتساع تقلصه. عند قوة معينة (مثالية) ، يصبح السعة القصوى. ومع ذلك ، إذا زادت قوة التحفيز بشكل أكبر ، فإن اتساع الانكماش لا يزداد بل ينخفض بسبب الاكتئاب الكاثودي. مثل هذه القوة ستكون متشائمة. يفسر رد فعل العضلات هذا بحقيقة أنها تتكون من ألياف ذات استثارة مختلفة ، وبالتالي ، فإن زيادة قوة التهيج مصحوبة بإثارة عدد متزايد منها. في القوة المثلى ، تشارك جميع الألياف في الانكماش. الاكتئاب الكاثودي هو انخفاض في الاستثارة تحت تأثير تيار مزيل للاستقطاب - كاثود ، أو قوة كبيرة أو مدة.
^ أوضاع التخفيض. القوة وعمل العضلات.
هناك الأنماط التالية لتقلص العضلات:
1. تقلصات متساوية التوتر. يتناقص طول العضلة ، لكن النغمة لا تتغير. لا يشاركون في الوظائف الحركية للجسم.
2-الانكماش الأحادي. لا يتغير طول العضلة ، بل تزداد النغمة. العمل الساكن الأساسي ، مثل الحفاظ على وضعية الجسم.
3. الانقباضات السمعية. كما يتغير طول ونغمة العضلات. بمساعدة منهم ، حركة الجسم ، تحدث أعمال حركية أخرى.
القوة العضلية القصوى هي مقدار التوتر الأقصى الذي يمكن أن تتطور إليه العضلة. يعتمد ذلك على بنية العضلات وحالتها الوظيفية وطولها الأولي والجنس والعمر ودرجة تدريب الشخص.
اعتمادًا على الهيكل ، توجد عضلات ذات ألياف متوازية (على سبيل المثال ، خياط) ، مغزلي (العضلة ذات الرأسين العضدية) ، ريشية (العجل). هذه الأنواع من العضلات لها منطقة مقطعية فسيولوجية مختلفة. إنه مجموع مناطق المقطع العرضي لجميع ألياف العضلات التي تتكون منها العضلة. أكبر منطقة مستعرضة فسيولوجية ، وبالتالي القوة ، هي في عضلات البينيت. أصغر العضلات بترتيب متوازي من الألياف (الشكل).
مع التمدد المعتدل للعضلة ، تزداد قوة تقلصها ، ولكن مع التمدد المفرط تقل. مع التسخين المعتدل ، يزداد أيضًا وينخفض مع التبريد. تقل قوة العضلات مع التعب واضطرابات التمثيل الغذائي وما إلى ذلك. يتم تحديد القوة القصوى لمجموعات العضلات المختلفة بواسطة مقاييس الدينامومتر والمعصم والرافعة المميتة وما إلى ذلك.
لمقارنة قوى العضلات المختلفة ، يتم تحديد قوتها المحددة أو المطلقة. إنه يساوي الحد الأقصى مقسومًا على مربع. رؤية منطقة المقطع العرضي للعضلة. القوة المحددة لعضلة الساق البشرية هي 6.2 كجم / سم 2 ، والعضلة ثلاثية الرؤوس 16.8 كجم / سم 2 ، وعضلة المضغ 10 كجم / سم 2.
ينقسم عمل العضلات إلى ديناميكي وثابت ، ويتم تنفيذ الديناميكي عند تحريك الحمولة. أثناء العمل الديناميكي ، يتغير طول العضلة وتوترها. لذلك ، تعمل العضلات في وضع auxotonic. في عمل ثابتلا توجد حركة للبضائع ، أي تعمل العضلات في وضع متساوي القياس. العمل الديناميكي يساوي ناتج وزن الحمولة وارتفاع ارتفاعها أو مقدار تقصير العضلات (A = P * h). يقاس العمل بالكيلو جرام.م ، جول. يخضع اعتماد مقدار العمل على الحمل لقانون متوسط الأحمال. عندما يزداد الحمل ، يزداد عمل العضلات في البداية. في الأحمال المتوسطة ، يصبح الحد الأقصى. إذا استمرت الزيادة في الحمل ، فسيقل العمل (الشكل). نفس التأثير على حجم العمل له إيقاعه. يتم تنفيذ أقصى قدر من عمل العضلات بإيقاع متوسط. من الأهمية بمكان في حساب حجم عبء العمل تعريف قوة العضلات. هذا هو العمل المنجز لكل وحدة زمنية (P = A * T). الثلاثاء
^ تعب عضلي
التعب هو انخفاض مؤقت في أداء العضلات نتيجة العمل. يمكن أن يكون سبب إجهاد العضلة المعزولة بسبب تحفيزها الإيقاعي. نتيجة لذلك ، تقل قوة الانقباضات تدريجياً (الشكل). كلما زاد التردد ، وقوة التهيج ، وحجم الحمل ، كلما تطور التعب بشكل أسرع. مع التعب ، يتغير منحنى الانكماش الفردي بشكل كبير. تزداد مدة الفترة الكامنة ، وفترة التقصير ، وخاصة فترة الاسترخاء ، لكن السعة تتناقص (الشكل). كلما كان إجهاد العضلات أقوى ، زادت مدة هذه الفترات. في بعض الحالات ، لا يحدث الاسترخاء التام. تطور التقلص. هذه حالة من انقباض العضلات اللاإرادي المطول. يتم فحص عمل العضلات والتعب باستخدام التصوير الشعاعي.
في القرن الماضي ، بناءً على تجارب مع عضلات معزولة ، تم اقتراح 3 نظريات حول إجهاد العضلات.
1. نظرية شيف: التعب هو نتيجة لاستنفاد احتياطيات الطاقة في العضلات.
2. نظرية بفلوجر: التعب ناتج عن تراكم نواتج التمثيل الغذائي في العضلات.
3. نظرية فيروورن: التعب ناتج عن نقص الأكسجين في العضلات.
وبالفعل ، فإن هذه العوامل تساهم في الإرهاق في التجارب على العضلات المعزولة. يتم إزعاج إعادة تخليق ATP فيها ، وتتراكم أحماض اللاكتيك والبيروفيك ، ويكون محتوى الأكسجين غير كافٍ. ومع ذلك ، في الجسم ، تتلقى العضلات العاملة بشكل مكثف الأكسجين والعناصر الغذائية الضرورية ويتم إطلاقها من المستقلبات بسبب زيادة الدورة الدموية العامة والإقليمية. لذلك ، تم اقتراح نظريات أخرى عن التعب. على وجه الخصوص ، تلعب المشابك العصبية العضلية دورًا معينًا في التعب. يتطور التعب في المشبك بسبب استنفاد مخازن الناقلات العصبية. ومع ذلك ، فإن الدور الرئيسي في إجهاد الجهاز الحركي ينتمي إلى المراكز الحركية للجهاز العصبي المركزي. في القرن الماضي ، أثبت I.M. Sechenov أنه في حالة إرهاق عضلات إحدى اليدين ، يتم استعادة أدائها بشكل أسرع عند العمل باليد الأخرى أو القدمين. وأعرب عن اعتقاده أن هذا يرجع إلى تحول عمليات الإثارة من مركز محرك إلى آخر. ودعا الباقي مع ضم مجموعات عضلية أخرى نشطة. لقد ثبت الآن أن التعب الحركي مرتبط بتثبيط المراكز العصبية المقابلة ، نتيجة لعمليات التمثيل الغذائي في الخلايا العصبية ، وتدهور تركيب الناقلات العصبية ، وتثبيط الانتقال المتشابك.
^ وحدات المحرك
العنصر المورفو الوظيفي الرئيسي للجهاز العصبي العضلي للعضلات الهيكلية هو الوحدة الحركية (MU). ويشمل العصبون الحركي للنخاع الشوكي مع ألياف عضلية يغذيها محوره. داخل العضلة ، يشكل هذا المحور عدة فروع طرفية. يشكل كل فرع من هذه الفروع جهة اتصال - مشابك عصبية عضلية على ألياف عضلية منفصلة. تسبب النبضات العصبية القادمة من الخلايا العصبية الحركية تقلصات لمجموعة معينة من ألياف العضلات. تحتوي الوحدات الحركية للعضلات الصغيرة التي تؤدي حركات دقيقة (عضلات العين واليد) على كمية صغيرة من ألياف العضلات. في الحالات الكبيرة ، هناك مئات المرات أكثر. يعتمد كل DU على الميزات الوظيفيةتنقسم إلى 3 مجموعات:
I. بطيئة الدؤوبة. تتشكل من ألياف عضلية "حمراء" ، حيث يوجد عدد أقل من اللييفات العضلية. معدل تقلص وقوة هذه الألياف صغير نسبيًا ، لكنها ليست مرهقة للغاية. لذلك ، يشار إليها باسم منشط. يتم تنظيم تقلصات هذه الألياف بواسطة عدد صغير من الخلايا العصبية الحركية ، والتي تحتوي محاورها على عدد قليل من الفروع الطرفية. مثال على ذلك هو عضلة النعل.
IIB. سريع وسهل التعب. تحتوي ألياف العضلات على العديد من اللييفات العضلية وتسمى "بيضاء". يتعاقد بسرعة ويطور قوة كبيرة ، لكن يتعب بسرعة. لذلك ، يطلق عليهم المرحلة. الخلايا العصبية الحركية لهذه MUs هي الأكبر ، ولها محور عصبي سميك مع العديد من الفروع الطرفية. يولدون نبضات عصبية عالية التردد. عضلات العين.
IIA. سريع ومقاوم للتعب. يشغلون منصب وسيط.
^ فسيولوجيا العضلات الملساء
توجد العضلات الملساء في جدران معظم أعضاء الجهاز الهضمي ، والأوعية الدموية ، والقنوات الإخراجية من الغدد المختلفة ، والجهاز البولي. فهي لا إرادية وتوفر التمعج للجهاز الهضمي والبولي ، مما يحافظ على توتر الأوعية الدموية. على عكس الهيكل العظمي ، تتكون العضلات الملساء من خلايا غالبًا على شكل مغزل وصغيرة الحجم ، والتي لا تحتوي على خطوط عرضية. هذا الأخير يرجع إلى حقيقة أن الجهاز المقلص لا يحتوي على هيكل مرتب. تتكون اللييفات العضلية من خيوط رقيقة من الأكتين تعمل في اتجاهات مختلفة وتلتصق بأجزاء مختلفة من غمد الليف العضلي. توجد الألياف الأولية للميوسين بجوار الأكتين. لا تشكل عناصر الشبكة الساركوبلازمية نظامًا من الأنابيب. ترتبط خلايا العضلات المنفصلة ببعضها البعض عن طريق ملامسات ذات مقاومة كهربائية منخفضة - الروابط ، مما يضمن انتشار الإثارة في جميع أنحاء بنية العضلات الملساء. استثارة وتوصيل العضلات الملساء أقل من تلك الموجودة في الهيكل العظمي.
تبلغ إمكانات الغشاء 40-60 مللي فولت ، نظرًا لأن غشاء MMC له نفاذية عالية نسبيًا لأيونات الصوديوم. علاوة على ذلك ، في العديد من العضلات الملساء ، لا يكون MP ثابتًا. يتناقص بشكل دوري ويعود مرة أخرى إلى مستواه الأصلي. تسمى هذه التذبذبات الموجات البطيئة (SW). عندما يصل الجزء العلوي من الموجة البطيئة إلى مستوى حرج من إزالة الاستقطاب ، تبدأ إمكانات الفعل في الظهور عليها ، مصحوبة بانقباضات (الشكل). يتم إجراء MV و PD من خلال عضلات ملساء بسرعة 5 إلى 50 سم / ثانية فقط. تسمى هذه العضلات الملساء نشطة تلقائيًا ، أي هم تلقائي. على سبيل المثال ، بسبب هذا النشاط ، يحدث التمعج المعوي. توجد أجهزة تنظيم ضربات القلب في التمعج المعوي في الأقسام الأولية من الأمعاء المقابلة.
يرجع توليد AP في SMCs إلى دخول أيونات الكالسيوم فيها. تختلف آليات الاقتران الكهروميكانيكي أيضًا. يتطور التقلص بسبب دخول الكالسيوم إلى الخلية أثناء PD ، وأهم بروتين خلوي ، كالموديولين ، يتوسط علاقة الكالسيوم بقصر اللييفات العضلية.
منحنى الانكماش مختلف أيضًا. الفترة الكامنة ، وهي فترة التقصير ، وخاصة الاسترخاء ، أطول بكثير من فترة عضلات الهيكل العظمي. يستمر الانكماش لبضع ثوان. تتميز العضلات الملساء ، على عكس العضلات الهيكلية ، بظاهرة النغمة البلاستيكية. هذه القدرة في حالة انخفاض لفترة طويلة دون استهلاك كبير للطاقة والتعب. بفضل هذه الخاصية ، يتم الحفاظ على شكل الأعضاء الداخلية ونغمة الأوعية الدموية. بالإضافة إلى ذلك ، فإن خلايا العضلات الملساء نفسها هي مستقبلات تمدد. عندما يتم تمديدها ، يبدأ إنشاء نقاط الوصول ، مما يؤدي إلى انخفاض في SMC. تسمى هذه الظاهرة الآلية العضلية لتنظيم نشاط الانقباض.