Какво е клетъчна мембрана и нейните функции. Историята на откриването и изследването на клетъчната мембрана. Плазмена мембрана, структура, функции
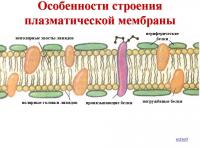
Отвън клетката е покрита с плазмена мембрана (или външна клетъчна мембрана) с дебелина около 6-10 nm.
Клетъчната мембрана е плътен филм от протеини и липиди (главно фосфолипиди). Липидните молекули са разположени подредено - перпендикулярно на повърхността, в два слоя, така че частите им, които интензивно взаимодействат с водата (хидрофилни), са насочени навън, а частите, които са инертни към водата (хидрофобни), са насочени навътре.
Протеиновите молекули са разположени в непрекъснат слой върху повърхността на липидната рамка от двете страни. Някои от тях са потопени в липидния слой, а други преминават през него, образувайки зони, пропускливи за вода. Тези протеини правят различни функции- някои от тях са ензими, други са транспортни протеини, участващи в преноса на определени вещества от околната среда в цитоплазмата и в обратна посока.
Основни функции на клетъчната мембрана
Едно от основните свойства на биологичните мембрани е селективната пропускливост (полупропускливост)- някои вещества преминават през тях трудно, други лесно и дори към по-висока концентрация.Така че за повечето клетки концентрацията на Na йони вътре е много по-ниска отколкото в заобикаляща среда. За K йони е характерно обратното съотношение: тяхната концентрация вътре в клетката е по-висока, отколкото извън нея. Следователно Na йоните винаги се стремят да влязат в клетката, а K йоните - да излязат навън. Изравняването на концентрациите на тези йони се предотвратява от наличието в мембраната на специална система, която играе ролята на помпа, която изпомпва Na йони от клетката и едновременно с това изпомпва K йони вътре.
Желанието на Na йоните да се движат отвън навътре се използва за транспортиране на захари и аминокиселини в клетката. С активното отстраняване на Na йони от клетката се създават условия за навлизане в нея на глюкоза и аминокиселини.
В много клетки абсорбцията на вещества се извършва също чрез фагоцитоза и пиноцитоза. При фагоцитозагъвкавата външна мембрана образува малка вдлъбнатина, където влиза уловената частица. Тази вдлъбнатина се увеличава и, заобиколена от част от външната мембрана, частицата се потапя в цитоплазмата на клетката. Феноменът на фагоцитоза е характерен за амеба и някои други протозои, както и за левкоцити (фагоцити). По същия начин има абсорбция от клетките на течности, съдържащи необходими на клеткатавещества. Това явление е наречено пиноцитоза.
Външните мембрани на различните клетки се различават значително както по химичния състав на техните протеини и липиди, така и по относителното им съдържание. Именно тези особености определят разнообразието във физиологичната активност на мембраните на различните клетки и тяхната роля в живота на клетките и тъканите.
Ендоплазменият ретикулум на клетката е свързан с външната мембрана. С помощта на външни мембрани, Различни видовемеждуклетъчни контакти, т.е. комуникация между отделните клетки.
Много видове клетки се характеризират с наличието на тяхната повърхност Голям бройиздатини, гънки, микровили. Те допринасят както за значително увеличаване на повърхността на клетките и подобряване на метаболизма, така и за по-силни връзки на отделните клетки една с друга.
От външната страна на клетъчната мембрана растителните клетки имат дебели мембрани, които са ясно видими в оптичен микроскоп, състоящи се от целулоза (целулоза). Те създават здрава опора за растителните тъкани (дърво).
Някои клетки от животински произход също имат редица външни структури, които са разположени върху клетъчната мембрана и имат защитен характер. Пример за това е хитинът на покривните клетки на насекомите.
Функции на клетъчната мембрана (накратко)
| функция | Описание |
|---|---|
| защитна бариера | Разделя вътрешните органели на клетката от външна среда |
| Регулаторен | Той регулира обмена на вещества между вътрешното съдържание на клетката и външната среда. |
| Разграничаване (компартментализация) | Разделяне на вътрешното пространство на клетката на независими блокове (отделения) |
| Енергия | - Натрупване и трансформиране на енергия; - светлинни реакции на фотосинтезата в хлоропластите; - Абсорбция и секреция. |
| Рецептор (информация) | Участва в образуването на възбуждане и неговото провеждане. |
| Мотор | Осъществява движението на клетката или отделните й части. |
Основни клетъчни мембрани:
плазмената мембрана
Плазмената мембрана, обграждаща всяка клетка, определя нейния размер, осигурява транспорта на малки и големи молекули от клетката и в клетката и поддържа разликата в концентрациите на йони от двете страни на мембраната. Мембраната участва в междуклетъчните контакти, възприема, усилва и предава сигнали от външната среда в клетката. Мембраната е свързана с много ензими, които катализират биохимичните реакции.
ядрена мембрана
Ядрената обвивка се състои от външна и вътрешна ядрена мембрана. Ядрената мембрана има пори, през които РНК проникват от ядрото в цитоплазмата, а регулаторните протеини от цитоплазмата в ядрото.
Вътрешната ядрена мембрана съдържа специфични протеини, които имат места за свързване на основните полипептиди на ядрената матрица - ламин А, ламин В и ламин С. Важна функция на тези протеини е разпадането на ядрената мембрана по време на митоза.
Мембрана на ендоплазмения ретикулум (ER).
ER мембраната има множество гънки и гънки. Той образува непрекъсната повърхност, която ограничава вътрешното пространство, наречено ER кухина. Грубият ER е свързан с рибозоми, върху които се синтезират протеини на плазмената мембрана, ER, апарат на Голджи, лизозоми и секретирани протеини. Регионите на ER, които не съдържат рибозоми, се наричат гладки ER. Тук последният етап от биосинтезата на холестерола, фосфолипидите, реакцията на окисление на собствените метаболити и чужди вещества с участието на мембранни ензими - цитохром Р 450, цитохром Р 450 редуктаза, цитохром b 5 редуктаза и цитохром b 5
апарат на Голджи
Апаратът на Голджи е важен мембранен органел, отговорен за модификацията, натрупването, сортирането и насочването на различни вещества в съответните вътреклетъчни отделения, както и извън клетката. Специфични ензими на мембраната на комплекса на Голджи, гликозилтрансфераза, гликозилиращи протеини в остатъците от серин, треонин или амидна група на аспарагин, завършват образуването на сложни протеини - гликопротеини.
Митохондриални мембрани
Митохондриите са органели с двойна мембрана, специализирани в синтеза на АТФ чрез окислително фосфорилиране. Отличителна черта на външната митохондриална мембрана е съдържанието на голямо количество поринов протеин, който образува пори в мембраната. Благодарение на порина, външната мембрана е свободно пропусклива за неорганични йони, метаболити и дори малки протеинови молекули (по-малко от 10 kD). За големите протеини външната мембрана е непропусклива, което позволява на митохондриите да предпазват протеините в междумембранното пространство от изтичане в цитозола.
Вътрешната мембрана на митохондриите се характеризира с високо съдържание на протеини, около 70%, които изпълняват главно каталитични и транспортни функции. Мембранните транслокази осигуряват селективен трансфер на вещества от междумембранното пространство към матрицата и обратно; ензимите участват в електронния транспорт (електронно-транспортна верига) и синтеза на АТФ.
Мембрана на лизозомите
Лизозомната мембрана играе ролята на "щит" между активните ензими (повече от 50), които осигуряват реакции за разграждане на протеини, въглехидрати, мазнини, нуклеинова киселина, и останалото съдържание на клетката. Мембраната съдържа уникални протеини, например АТФ-зависима протонна помпа (помпа), която поддържа кисела среда (рН 5), необходима за действието на хидролитичните ензими (протеази, липази), както и транспортни протеини, които позволяват разграждането на макромолекулите продукти да напуснат лизозомата. Такива мембрани ги предпазват от действието на протеазите.
Общи функции на биологичните мембрани следното:
Те разграничават съдържанието на клетката от външната среда и съдържанието на органелите от цитоплазмата.
Те осигуряват транспорт на вещества в и извън клетката, от цитоплазмата към органелите и обратно.
Те играят ролята на рецептори (приемане и преобразуване на сигнали от околната среда, разпознаване на клетъчни вещества и др.).
Те са катализатори (осигуряващи мембранни химични процеси).
Участвайте в трансформацията на енергията.
Общи свойства на биологичните мембрани
Всички без изключение клетъчни мембрани са изградени според общ принцип: това са тънки липопротеинови филми, състоящи се от двоен слой липидни молекули, в които са включени протеинови молекули. В тегловно отношение, в зависимост от вида на мембраните, липидите представляват 25-60%, а протеините - 40-75%. Много мембрани съдържат въглехидрати, чието количество може да достигне 2-10%.
Биологичните мембрани много селективно пропускат вещества от околния разтвор. Те пропускат вода доста лесно и улавят повечето от веществата, разтворими във вода, и предимно йонизирани вещества или такива, които носят електрически заряд. Поради това биомембраните са добри електрически изолатори в солеви разтвори.
Мембранна подложка е двоен липиден слойв образуването на които участват фосфолипиди и гликолипиди. Липидният бислой се образува от два реда липиди, чиито хидрофобни радикали са скрити вътре, а хидрофилните групи са обърнати навън и са в контакт с водната среда. Протеиновите молекули са сякаш "разтворени" в липидния двоен слой
Напречно сечение на плазмената мембрана
Липиден състав на мембраните:
Фосфолипиди.Всички фосфолипиди могат да бъдат разделени на 2 групи - глицерофосфолипиди и сфингофосфолипиди. Глицерофосфолипидите се класифицират като производни на фосфатидната киселина. Най-често срещаните мембранни глицерофосфолипиди са фосфатидилхолини и фосфатидилетаноламини. Огромен брой различни фосфолипиди са открити в мембраните на еукариотните клетки и те са разпределени неравномерно върху различните клетъчни мембрани. Тази неравномерност се отнася до разпределението както на полярните "глави", така и на ацилните остатъци.
Специфични фосфолипиди на вътрешната мембрана на митохондриите са кардиолипините (дифосфатидилглицероли), изградени на базата на глицерол и два остатъка от фосфатидна киселина. Те се синтезират от ензими на вътрешната митохондриална мембрана и съставляват около 22% от всички мембранни фосфолипиди.
Плазмените мембрани на клетките съдържат сфингомиелини в значителни количества. Сфингомиелините са изградени на основата на церамид, ацилиран аминоалкохол на сфингозин. Полярната група се състои от остатък от фосфорна киселина и холин, етаноламин или серин. Сфингомиелините са основните липиди в миелиновата обвивка на нервните влакна.
Гликолипиди.В гликолипидите хидрофобната част е представена от керамид. Хидрофилна група - въглехидратен остатък, свързан чрез гликозидна връзка към хидроксилната група при първия въглероден атом на церамида. В зависимост от дължината и структурата на въглехидратната част има цереброзиди,съдържащи моно- или олигозахариден остатък, и ганглиозиди,към ОН групата на който е прикрепен сложен, разклонен олигозахарид, съдържащ N-ацетилневраминова киселина (NANA).
Полярните "глави" на гликосфинголипидите са разположени на външната повърхност на плазмените мембрани. Значителни количества гликолипиди се намират в мембраните на мозъчните клетки, еритроцитите и епителните клетки. Ганглиозидите на еритроцитите на различни индивиди се различават по структурата на олигозахаридните вериги, които проявяват антигенни свойства.
Холестерол.Холестеролът присъства във всички мембрани на животински клетки. Молекулата му се състои от твърдо хидрофобно ядро и гъвкава въглеводородна верига, единствената хидроксилна група е "полярната глава".
За животинска клетка средното моларно съотношение холестерол/фосфолипид е 0,3-0,4, но в плазмената мембрана това съотношение е много по-високо (0,8-0,9). Наличието на холестерол в мембраните намалява подвижността мастни киселини, намалява страничната дифузия на липидите и протеините и следователно може да повлияе на функциите на: мембранните протеини.
В растителните мембрани няма холестерол, но има растителни стероиди - ситостерол и стигмастерол.
Мембранни протеини: Прието е да се разделят на интегрални (трансмембранни) и периферни. Интеграл протеините имат обширни хидрофобни области на повърхността си и са неразтворими в вода. Те са свързани с мембранните липиди чрез хидрофобни взаимодействия и частично са потопени в дебелината на липидния двоен слой и често проникват в двойния слой, напускайки повърхностите са относително малки хидрофилни зони. Отделете тези протеини от мембраните могат да бъдат постигнати само с детергенти като додецилсулфат или соли жлъчни киселини, които разрушават липидния слой и превръщат протеина в разтворим форма (разтворете го) чрез образуване на асоциирани с него. Всички по-нататъшни операции Пречистването на интегралните протеини също се извършва в присъствието на детергенти. Периферните протеини са свързани с повърхността на липидния двоен слой електростатични сили и може да се отмие от мембраната със солеви разтвори.
23. Механизми за пренос на вещества през мембрани: проста дифузия, пасивен симпорт и антипорт, първичен активен транспорт, вторичен активен транспорт, регулирани канали (примери). Пренасяне през мембраната на макромолекули и частици. Участие на мембраните в междуклетъчните взаимодействия.
Има няколко механизми за транспортиране на веществата през мембраната .
дифузия- проникване на вещества през мембраната по концентрационния градиент (от областта, където концентрацията им е по-висока, към зоната, където концентрацията им е по-ниска). Дифузният транспорт на вещества (вода, йони) се осъществява с участието на мембранни протеини, които имат молекулни пори, или с участието на липидната фаза (за мастноразтворимите вещества).
С улеснена дифузияспециални мембранни носители протеини селективно се свързват с един или друг йон или молекула и ги пренасят през мембраната по концентрационен градиент.
Улеснена дифузия на вещества
Транслоказните протеини съществуват в клетъчните мембрани. Взаимодействайки със специфичен лиганд, те осигуряват неговата дифузия (транспорт от зона с по-висока концентрация до област с по-ниска концентрация) през мембраната. За разлика от протеиновите канали, транслоказите претърпяват конформационни промени в процеса на взаимодействие с лиганда и пренасянето му през мембраната. Кинетично, преносът на вещества чрез улеснена дифузия наподобява ензимна реакция. За транслоказите има насищаща концентрация на лиганда, при която всички места на свързване на протеина с лиганда са заети и протеините работят с максимална скорост Vmax. Следователно скоростта на транспортиране на веществата чрез улеснена дифузия зависи не само от концентрационния градиент на транспортирания лиганд, но и от броя на протеините носители в мембраната.
Има транслокази, които пренасят само едно водоразтворимо вещество от едната страна на мембраната до другата. Такъв прост транспорт се нарича "пасивен унипорт". Пример за унипорт е функционирането на GLUT-1, транслоказа, която транспортира глюкоза през еритроцитната мембрана:
Улеснена дифузия (uniport) на глюкоза в еритроцитите с помощта на GLUT-1 (S - глюкозна молекула). Молекулата на глюкозата е свързана с носител върху външната повърхност на плазмената мембрана. Настъпва конформационна промяна и центърът на носителя, зает от глюкоза, е изложен на вътрешността на клетката. Поради конформационни промени носителят губи своя афинитет към глюкозата и молекулата се освобождава в цитозола на клетката. Отделянето на глюкозата от носителя причинява конформационна промяна в протеина и той се връща към първоначалната си "информация".
Някои транслокази могат да пренасят две различни вещества по концентрационен градиент в една и съща посока - пасивен симпорт , или в противоположни посоки - пасивен антипорт .
Пример за транслоказа, работеща чрез пасивен антипортов механизъм, е анионният транспортер на еритроцитната мембрана. Вътрешната митохондриална мембрана съдържа много транслокази, които извършват пасивен антипорт. В процеса на такъв трансфер се извършва еквивалентен обмен на йони, но не винаги еквивалентен обмен на заряд.
първичен активен транспорт
Транспортът на някои неорганични йони протича срещу концентрационния градиент с участието на транспортни АТФази (йонни помпи). Всички йонни помпи едновременно служат като ензими, способни на автофосфорилиране и автодефосфорилиране. АТФазите се различават по йонна специфичност, брой пренесени йони и посока на транспорт. В резултат на функционирането на АТФ-аза, транспортираните йони се натрупват от едната страна на мембраната. Ma+,K+-ATPase, Ca2+-ATPase и H+,K+,-ATPase на стомашната лигавица са най-разпространени в плазмената мембрана на човешките клетки.
Na+, K+-АТФаза
Този ензим носител катализира АТФ-зависимия транспорт на Na+ и K+ йони през плазмената мембрана. Ka+,K+-ATPase се състои от α и β субединици; α - каталитична голяма субединица, и β - малка субединица (гликопротеин). Активната форма на транслоказата е тетрамерът (αβ)2.
Na+,K+-ATPase е отговорна за поддържането на висока концентрация на K+ в клетката и ниска концентрация на Na+. Тъй като Na + D + -ATPase изпомпва три положително заредени йона и изпомпва два, върху мембраната възниква електрически потенциал с отрицателна стойност от вътрешната страна на клетката по отношение на нейната външна повърхност.
Са2+-АТФазалокализиран не само в плазмената мембрана, но и в ER мембраната. Ензимът се състои от десет трансмембранни домена, обхващащи клетъчната мембрана. Между втория и третия домен има няколко остатъка от аспарагинова киселина, участващи в свързването на калций. Регионът между четвъртия и петия домен има център за прикрепване на АТФ и автофосфорилиране в остатъка от аспарагинова киселина. Са2+-АТФазите на плазмените мембрани на някои клетки се регулират от протеина калмодулин. Всяка от Са2+-АТФазите на плазмената мембрана и ER е представена от няколко изоформи.
вторичен активен транспорт
Преносът на някои разтворени вещества срещу концентрационния градиент зависи от едновременния или последователен транспорт на друго вещество по концентрационния градиент в същата посока (активен симпорт) или в обратната посока (активен антипорт). В човешките клетки Na+ е най-често йонът, транспортиран по концентрационния градиент.

Последователност на събитията в процеса на работа на Са2*-АТФ-аза.
1 - свързване на два калциеви йона от АТФ-азно място, обърнато към цитозола;
2 - промяна в заряда и конформацията на ензима (АТФ-аза), причинена от добавянето на два Са2+ йона, води до повишаване на афинитета към АТФ и активиране на автофосфорилирането;
3 - автофосфорилирането е придружено от информационни промени, АТФазата се затваря с вътремембрана и се отваря отвън;
4 - има намаляване на афинитета на свързващите центрове за калциевите йони и те се отделят от АТФ-аза;
5 - автодефосфорилирането се активира от магнезиеви йони, в резултат на което Ca2+-ATP-азата губи фосфорен остатък и два Mg2+ йона;
6 - ATPase се връща в първоначалното си състояние.
Пример за този вид транспорт е Na+,Ca2+ обменникът на плазмената мембрана (активен антипорт), натриевите йони се транспортират в клетката по концентрационния градиент, а Ca2+ йоните излизат от клетката срещу концентрационния градиент.
Съгласно механизма на активния симпорт се извършва абсорбцията на глюкоза от чревните клетки и реабсорбцията на глюкоза и аминокиселини от първичната урина от бъбречните клетки.
Транспорт през мембраната на макромолекули и частици: ендоцитоза и екзоцитоза
Макромолекулите на протеините, нуклеиновите киселини, полизахаридите, липопротеиновите комплекси и др. не преминават през клетъчните мембрани, за разлика от йоните и мономерите. Транспортирането на макромолекули, техните комплекси и частици в клетката става по съвсем различен начин - чрез ендоцитоза. При ендоцитоза (ендо...- вътре) определен участък от плазмалемата улавя и, така да се каже, обгръща извънклетъчния материал, затваряйки го в мембранна вакуола, възникнала в резултат на мембранна инвагинация. Впоследствие такава вакуола е свързана с лизозома, чиито ензими разграждат макромолекулите до мономери.
Обратният процес на ендоцитоза екзоцитоза (екзо...- отвън). Благодарение на него клетката премахва вътреклетъчните продукти или неразградените остатъци, затворени във вакуоли или везикули. Везикулата се приближава до цитоплазмената мембрана, слива се с нея и съдържанието й се освобождава в околната среда. Как се отделят храносмилателни ензими, хормони, хемицелулоза и др.
По този начин биологичните мембрани, като основни структурни елементи на клетката, служат не само като физически граници, но и като динамични функционални повърхности. На мембраните на органелите се извършват множество биохимични процеси, като активна абсорбция на вещества, преобразуване на енергия, синтез на АТФ и др.
УЧАСТИЕ НА МЕМБРАНИТЕ В МЕЖДУКЛЕТЪЧНИТЕ ВЗАИМОДЕЙСТВИЯ
Плазмената мембрана на еукариотните клетки съдържа много специализирани рецептори, които, взаимодействайки с лиганди, предизвикват специфични клетъчни реакции. Някои рецептори свързват сигнални молекули - хормони, невротрансмитери, други - хранителни вещества и метаболити, а трети участват в клетъчната адхезия. Този клас включва рецептори, необходими за клетъчно разпознаване и адхезия, както и рецептори, отговорни за клетъчното свързване към протеини на извънклетъчния матрикс като фибронектин или колаген.
Способността на клетките за специфично взаимно разпознаване и адхезия е важна за ембрионалното развитие. При възрастни адхезивните взаимодействия клетка-клетка и клетка-матрикс продължават да бъдат от съществено значение за поддържане на стабилността на тъканта. В голямо семейство рецептори за клетъчна адхезия интегрините, селектините и кадхерините са най-изследваните.
Интегрини- обширно суперсемейство от хомоложни рецептори на клетъчната повърхност за извънклетъчни матрични молекули, като колаген, фибронектин, ламинин и др. Като трансмембранни протеини, те взаимодействат както с извънклетъчните молекули, така и с вътреклетъчните протеини на цитоскелета. Благодарение на това интегрините участват в преноса на информация от извънклетъчната среда към клетката, като по този начин определят посоката на нейната диференциация, форма, митотична активност и способност за миграция. Преносът на информация може да върви и в обратна посока – от вътреклетъчните протеини през рецептора към извънклетъчния матрикс.
Примери за някои интегрини:
рецептори за протеини на извънклетъчния матрикс. Те се свързват с гликопротеиновите компоненти на екстрацелуларния матрикс, по-специално фибронектин, ламинин и витронектин (вижте точка 15);
тромбоцитните интегрини (IIb и IIIa) участват в агрегацията на тромбоцитите, която възниква по време на кръвосъсирването;
левкоцитни адхезионни протеини. За да мигрират към мястото на инфекцията и възпалението, левкоцитите трябва да взаимодействат със съдовите ендотелни клетки. Това взаимодействие може да медиира свързването на Т-лимфоцитите с фибробластите по време на възпаление.
Кадхерини и селектинисемейства на трансмембранни Ca 2+ -зависими гликопротеини, участващи в междуклетъчната адхезия. Три възможни начиниучастие на рецептори от този тип в междуклетъчната адхезия.


фибронектинов рецептор.Фибронектиновият рецептор принадлежи към семейството на интегрините. Всяка субединица има единичен трансмембранен домен, къс цитоплазмен домен и разширен N-екстрацелуларен домен. И двете субединици (α, β) на интегрина са гликозилирани и държани заедно чрез нековалентни връзки, α-субединицата се синтезира като единична полипептидна верига, която след това се разцепва на малка трансмембранна верига и голяма извънклетъчна верига, свързана с дисулфид мостове. β-субединицата съдържа 4 повторения от по 40 аминокиселинни остатъка всеки. α-субединиците са богати на цистеин и съдържат много вътреверижни дисулфидни връзки (не са показани на фигурата). Чрез свързване с фибронектин отвън и с цитоскелета вътре в клетката, интегринът действа като трансмембранен линкер.
Методи за взаимодействие между повърхностните молекули на клетката в процеса на междуклетъчна адхезия.А - рецепторите на една клетка могат да се свързват със същите рецептори на съседни клетки (хомофилно свързване); B - рецепторите на една клетка могат да се свързват с рецептори на друг тип съседни клетки (хетерофилно свързване); В - рецепторите на клетъчната повърхност на съседни клетки могат да комуникират помежду си, използвайки поливалентни линкерни молекули.
Кадхерините от различни тъкани са много сходни, с 50-60% хомоложни аминокиселинни последователности. Всеки рецептор има един трансмембранен домен.
Най-пълно са характеризирани три групи кадхеринови рецептори:
Е-кадхеринът се намира на повърхността на много клетки в епителните и ембрионалните тъкани;
N-кадхеринът е локализиран на повърхността на нервните клетки, сърдечните клетки и лещата;
Р-кадхеринът се намира върху клетките на плацентата и епидермиса.
Кадхерините играят важна роля в първоначалната междуклетъчна адхезия, на етапите на морфо- и органогенезата и осигуряват структурната цялост и полярност на тъканите, особено на епителния монослой.
В семейството избирамрецептори, три протеина са най-добре проучени: L-селектин, P-селектин и E-селектин. Извънклетъчната част на селектините се състои от 3 домени: първият домен е представен от 2-9 блока от повтарящи се аминокиселинни остатъци (комплемент-регулаторен протеин), вторият е домейнът на епидермалния растежен фактор (EGF), третият е N-терминалният лектинов домен. Селектините L, P, E се различават по броя на блоковете в комплемент-регулаторния протеин. Лектините са семейство от протеини, които специфично взаимодействат с определени последователности от въглехидратни остатъци в гликопротеини, протеогликани и гликолипиди на извънклетъчния матрикс.
Кратко описание:
Сазонов В.Ф. 1_1 Структурата на клетъчната мембрана [Електронен ресурс] // Кинезиолог, 2009-2018: [уебсайт]. Дата на актуализация: 06.02.2018 г..__.201_). _Описана е структурата и функционирането на клетъчната мембрана (синоними: плазмалема, плазмолема, биомембрана, клетъчна мембрана, външна клетъчна мембрана, клетъчна мембрана, цитоплазмена мембрана). Тази първоначална информация е необходима както за цитологията, така и за разбиране на процесите нервна дейност: нервна възбуда, инхибиране, работа на синапсите и сензорните рецептори.
клетъчна мембрана (плазма Алема или плазма Олема)
Определение на понятието
Клетъчната мембрана (синоними: плазмалема, плазмолема, цитоплазмена мембрана, биомембрана) е тройна липопротеинова (т.е. "мастно-протеинова") мембрана, която отделя клетката от околната среда и осъществява контролиран обмен и комуникация между клетката и нейната среда.
Основното в това определение не е, че мембраната отделя клетката от околната среда, а просто тя свързва клетка с околната среда. Мембраната е активен структура на клетката, тя непрекъснато работи.
Биологичната мембрана е ултратънък бимолекулен филм от фосфолипиди, инкрустиран с протеини и полизахариди. Тази клетъчна структура е в основата на бариерните, механичните и матричните свойства на живия организъм (Антонов В.Ф., 1996).
Фигуративно представяне на мембраната
На мен клетъчната мембранаПредставен е като решетъчна ограда с много врати в нея, която огражда определена територия. Всякакви малки живи същества могат свободно да се движат напред-назад през тази ограда. Но по-големите посетители могат да влязат само през вратите и дори тогава не всички. Различните посетители имат ключове само за собствените си врати и не могат да минават през вратите на други хора. И така, през тази ограда има непрекъснати потоци от посетители напред-назад, т.к Главна функцияоградните мембрани са двойни: за да отделят територията от околното пространство и в същото време да я свържат с околното пространство. За това има много дупки и врати в оградата - !
Мембранни свойства
1. Пропускливост.
2. Полупропускливост (частична пропускливост).
3. Селективна (синоним: избирателна) пропускливост.
4. Активна пропускливост (синоним: активен транспорт).
5. Контролирана пропускливост.
Както можете да видите, основното свойство на мембраната е нейната пропускливост по отношение на различни вещества.
6. Фагоцитоза и пиноцитоза.
7. Екзоцитоза.
8. Наличието на електрически и химичен потенциал, по-точно потенциалната разлика между вътрешната и външната страна на мембраната. Образно може да се каже така "мембраната превръща клетката в "електрическа батерия" чрез контролиране на йонните потоци". подробности: .
9. Промени в електрическия и химичния потенциал.
10. Раздразнителност. Специални молекулни рецептори, разположени върху мембраната, могат да се свързват със сигнални (контролни) вещества, в резултат на което състоянието на мембраната и цялата клетка може да се промени. Молекулярните рецептори задействат био химична реакцияв отговор на комбинацията от лиганди (контролни вещества) с тях. Важно е да се отбележи, че сигналното вещество действа върху рецептора отвън, докато промените продължават вътре в клетката. Оказва се, че мембраната предава информация от околната среда към вътрешната среда на клетката.
11. Каталитична ензимна активност. Ензимите могат да бъдат вградени в мембраната или свързани с нейната повърхност (както вътре, така и извън клетката) и там те извършват своята ензимна активност.
12. Промяна на формата на повърхността и нейната площ. Това позволява на мембраната да образува израстъци навън или, обратно, инвагинации в клетката.
13. Способност за контакт с други клетъчни мембрани.
14. Адхезия - способността за залепване към твърди повърхности.
Кратък списък на свойствата на мембраната
- Пропускливост.
- Ендоцитоза, екзоцитоза, трансцитоза.
- потенциали.
- раздразнителност.
- ензимна активност.
- Контакти.
- Адхезия.
Функции на мембраната
1. Непълна изолация на вътрешното съдържание от външната среда.
2. Основното в работата на клетъчната мембрана е обмен различни вещества между клетката и извънклетъчната среда. Това се дължи на такова свойство на мембраната като пропускливост. В допълнение, мембраната регулира този обмен, като регулира пропускливостта си.
3. Друга важна функция на мембраната е създаване на разлика в химическия и електрическия потенциал между вътрешната и външната му страна. Поради това вътре в клетката има отрицателен електрически потенциал -.
4. През мембраната също се извършва обмен на информация между клетката и нейната среда. Специални молекулни рецептори, разположени върху мембраната, могат да се свързват с контролиращи вещества (хормони, медиатори, модулатори) и да задействат биохимични реакции в клетката, водещи до различни променив работата на клетката или в нейните структури.
Видео:Структурата на клетъчната мембрана
Видео лекция:Подробности за структурата на мембраната и транспорта
Структура на мембраната
Клетъчната мембрана има универсален трислоен структура. Средният му мастен слой е непрекъснат, а горният и долният протеинов слой го покриват под формата на мозайка от отделни протеинови области. Мастният слой е основата, която осигурява изолацията на клетката от околната среда, изолирайки я от околната среда. Сам по себе си той много зле пропуска водоразтворимите вещества, но лесно пропуска мастноразтворимите. Следователно, пропускливостта на мембраната за водоразтворими вещества (например йони) трябва да бъде осигурена със специални протеинови структури - и.
По-долу са дадени микроснимки на реални клетъчни мембрани на контактуващи клетки, получени с помощта на електронен микроскоп, както и схематичен чертеж, показващ трислойната мембрана и мозаечната природа на нейните протеинови слоеве. За да увеличите изображение, щракнете върху него.

Отделно изображение на вътрешния липиден (мастен) слой на клетъчната мембрана, пропит с интегрални вградени протеини. Горният и долният протеинов слой се отстраняват, за да не пречат на разглеждането на липидния двоен слой

Фигура по-горе: Непълно схематично представяне на клетъчната мембрана (клетъчна стена) от Wikipedia.
Обърнете внимание, че външният и вътрешният протеинов слой са отстранени от мембраната тук, така че можем да видим по-добре централния мастен двоен липиден слой. В истинската клетъчна мембрана големи протеинови "острови" плават отгоре и отдолу по мастния филм (малки топки на фигурата), а мембраната се оказва по-дебела, трислойна: протеин-мазнина-протеин . Така че всъщност е като сандвич от две протеинови "филии хляб" с дебел слой "масло" в средата, т.е. има трислойна структура, а не двуслойна.
На тази фигура малки сини и бели топчета съответстват на хидрофилните (намокряеми) „глави“ на липидите, а „струните“, прикрепени към тях, съответстват на хидрофобните (ненамокрящи се) „опашки“. От протеините са показани само интегрални мембранни протеини от край до край (червени глобули и жълти спирали). Жълтите овални точки вътре в мембраната са холестеролови молекули. Жълто-зелените вериги от перли от външната страна на мембраната са олигозахаридни вериги, които образуват гликокаликса. Гликокаликсът е като въглехидратен ("захарен") "пух" върху мембраната, образуван от дълги въглехидратно-протеинови молекули, стърчащи от нея.
Living е малка "протеиново-мазнина торба", пълна с полутечно желеобразно съдържание, което е проникнато от филми и тръби.
Стените на тази торбичка са изградени от двоен мастен (липиден) филм, покрит отвътре и отвън с протеини - клетъчната мембрана. Следователно се казва, че мембраната има трислойна структура : протеини-мазнини-протеини. Вътре в клетката също има много подобни мастни мембрани, които разделят вътрешното й пространство на отделения. Клетъчните органели са заобиколени от същите мембрани: ядро, митохондрии, хлоропласти. Така че мембраната е универсална молекулярна структура, присъща на всички клетки и всички живи организми.

Отляво - вече не реален, а изкуствен модел на парче от биологична мембрана: това е мигновена снимка на адипозен фосфолипиден двуслой (т.е. двоен слой) в процеса на неговото молекулярно динамично моделиране. Показана е изчислителната клетка на модела - 96 PQ молекули ( fосфатидил холин) и 2304 водни молекули, общо 20544 атома.
Вдясно е визуален модел на единична молекула от същия липид, от която е сглобен мембранният липиден двоен слой. Има хидрофилна (водолюбива) глава в горната част и две хидрофобни (страхуващи се от вода) опашки в долната част. Този липид има просто име: 1-стероил-2-докозахексаеноил-Sn-глицеро-3-фосфатидилхолин (18:0/22:6(n-3)cis PC), но не е необходимо да го запомняте, освен ако планирайте да накарате вашия учител да припадне от дълбочината на вашите знания.
Можете да дадете по-точно научно определение на клетка:
е подредена, структурирана хетерогенна система от биополимери, ограничена от активна мембрана, участваща в единен набор от метаболитни, енергийни и информационни процеси, а също така поддържаща и възпроизвеждаща цялата система като цяло.
Вътре в клетката също е проникнато от мембрани, а между мембраните няма вода, а вискозен гел / зол с променлива плътност. Следователно взаимодействащите молекули в клетката не се носят свободно, както в епруветка с воден разтвор, но предимно седят (имобилизирани) върху полимерните структури на цитоскелета или вътреклетъчните мембрани. И следователно химичните реакции протичат вътре в клетката почти като в твърдо тяло, а не в течност. Външната мембрана, която обгражда клетката, също е покрита с ензими и молекулярни рецептори, което я прави много активна част от клетката.
Клетъчната мембрана (плазмалема, плазмолема) е активна обвивка, която отделя клетката от околната среда и я свързва с околната среда. © Сазонов V.F., 2016.
От това определение за мембрана следва, че тя не просто ограничава клетката, но активно работещсвързвайки го с неговата среда.
Мазнината, която изгражда мембраните, е специална, така че нейните молекули обикновено се наричат не просто мазнини, но липиди, фосфолипиди, сфинголипиди. Мембранният филм е двоен, т.е. състои се от два залепени един за друг филма. Следователно в учебниците пише, че основата на клетъчната мембрана се състои от два липидни слоя (или " двуслоен", т.е. двоен слой). За всеки отделен липиден слой, едната страна може да бъде намокрена от вода, а другата не. Така че тези филми се залепват един с друг именно с техните немокрящи се страни.
бактериална мембрана
Обвивката на прокариотна клетка от грам-отрицателни бактерии се състои от няколко слоя, показани на фигурата по-долу.
Слоеве на черупката на грам-отрицателни бактерии:
1. Вътрешната трислойна цитоплазмена мембрана, която е в контакт с цитоплазмата.
2. Клетъчна стена, която се състои от муреин.
3. Външната трислойна цитоплазмена мембрана, която има същата система от липиди с протеинови комплекси като вътрешната мембрана.
Комуникацията на грам-отрицателните бактериални клетки с външния свят чрез такава сложна тристепенна структура не им дава предимство при оцеляване в тежки условия в сравнение с грам-положителните бактерии, които имат по-малко мощна обвивка. Те просто не го приемат добре високи температури, повишена киселинност и спадове на налягането.




Видео лекция: плазмената мембрана. Е.В. Cheval, Ph.D.
Видео лекция:Мембраната като клетъчна граница. А. Иляскин
Значение на мембранните йонни канали
Лесно е да се разбере, че само мастноразтворими вещества могат да навлязат в клетката през мембранния мастен филм. Това са мазнини, алкохоли, газове.Например в еритроцитите кислородът и въглеродният диоксид лесно преминават навътре и навън директно през мембраната. Но водата и водоразтворимите вещества (например йони) просто не могат да преминат през мембраната в никоя клетка. Това означава, че те се нуждаят от специални дупки. Но ако просто направите дупка в мастния филм, той веднага ще се стегне обратно. Какво да правя? В природата е намерено решение: необходимо е да се направят специални протеинови транспортни структури и да се разтегнат през мембраната. Така се получават каналите за преминаване на мастнонеразтворимите вещества – йонните канали на клетъчната мембрана.
И така, да дадете мембраната си допълнителни свойствапропускливост за полярни молекули (йони и вода), клетката синтезира специални протеини в цитоплазмата, които след това се интегрират в мембраната. Те са два вида: транспортни протеини (например транспортни АТФази) и каналообразуващи протеини (формиращи канали). Тези протеини са вградени в двойния мастен слой на мембраната и образуват транспортни структури под формата на транспортери или под формата на йонни канали. Различни водоразтворими вещества вече могат да преминават през тези транспортни структури, които иначе не могат да преминат през мастния мембранен филм.
Най-общо протеините, вградени в мембраната, също се наричат интегрална, именно защото те като че ли са включени в състава на мембраната и я проникват наскрізь. Други протеини, които не са интегрални, образуват, така да се каже, острови, които "плуват" на повърхността на мембраната: или по външната й повърхност, или по вътрешната. В крайна сметка всеки знае, че мазнината е добра смазка и лесно се плъзга по нея!
заключения
1. Като цяло мембраната е трислойна:
1) външният слой от протеинови "острови",
2) мастно двуслойно "море" (липиден двуслой), т.е. двоен липиден филм
3) вътрешния слой от протеинови "острови".
Но има и рехав външен слой - гликокаликс, който се образува от гликопротеини, стърчащи от мембраната. Те са молекулярни рецептори, към които се свързват сигналните контроли.
2. В мембраната са вградени специални протеинови структури, осигуряващи нейната пропускливост за йони или други вещества. Не трябва да забравяме, че на някои места морето от мазнини е пронизано с интегрални протеини. И именно интегралните протеини образуват специални транспортни структури клетъчна мембрана (вижте раздел 1_2 Мембранни транспортни механизми). Чрез тях веществата навлизат в клетката и също се извеждат от клетката навън.
3. Ензимните протеини могат да бъдат разположени от всяка страна на мембраната (външна и вътрешна), както и вътре в мембраната, които влияят както на състоянието на самата мембрана, така и на живота на цялата клетка.
Така че клетъчната мембрана е активна променлива структура, която активно работи в интерес на цялата клетка и я свързва с външния свят, а не е просто "защитна обвивка". Това е най-важното нещо, което трябва да знаете за клетъчната мембрана.
В медицината мембранните протеини често се използват като „мишени“ за лекарства. Като такива мишени действат рецептори, йонни канали, ензими, транспортни системи. Напоследък освен мембраната целта за лекарствени веществасъщо стават гени, скрити в клетъчното ядро.
Видео:Въведение в биофизиката на клетъчната мембрана: Структура на мембрана 1 (Владимиров Ю.А.)
Видео:История, структура и функции на клетъчната мембрана: Структура на мембраните 2 (Владимиров Ю.А.)
© 2010-2018 Сазонов V.F., © 2010-2016 kineziolog.bodhy.
Основен структурна единицажив организъм - клетка, която е диференцирана област от цитоплазмата, заобиколена от клетъчна мембрана. С оглед на факта, че клетката изпълнява много важни функции, като възпроизводство, хранене, движение, черупката трябва да бъде пластична и плътна.
История на откриването и изследването на клетъчната мембрана
През 1925 г. Грендел и Гордър правят успешен експеримент за идентифициране на "сенките" на еритроцитите или празните черупки. Въпреки няколко груби грешки, учените откриха липидния двоен слой. Тяхната работа е продължена от Даниели, Доусън през 1935 г., Робъртсън през 1960 г. В резултат на дългогодишна работа и натрупване на аргументи през 1972 г. Сингър и Никълсън създават флуиден мозаечен модел на структурата на мембраната. По-нататъшни експерименти и проучвания потвърдиха работата на учените.
Значение
Какво е клетъчна мембрана? Тази дума започва да се използва преди повече от сто години, в превод от латински означава "филм", "кожа". Така се обозначава границата на клетката, която е естествена бариера между вътрешното съдържание и външната среда. Структурата на клетъчната мембрана предполага полупропускливост, поради което влагата и хранителните вещества и продуктите на гниене могат свободно да преминават през нея. Тази обвивка може да се нарече основният структурен компонент на организацията на клетката.

Помислете за основните функции на клетъчната мембрана
1. Разделя вътрешното съдържание на клетката и компонентите на външната среда.
2. Спомага за поддържането на постоянен химичен състав на клетката.
3. Регулира правилния метаболизъм.
4. Осигурява взаимовръзка между клетките.
5. Разпознава сигнали.
6. Защитна функция.
"плазмена обвивка"
Външната клетъчна мембрана, наричана още плазмена мембрана, е ултрамикроскопичен филм с дебелина от пет до седем нанометра. Състои се главно от протеинови съединения, фосфолиди, вода. Филмът е еластичен, лесно абсорбира вода и също така бързо възстановява целостта си след повреда.
Различава се в универсална структура. Тази мембрана заема гранична позиция, участва в процеса на селективна пропускливост, екскреция на продуктите на гниене, синтезира ги. отношения със съседите и надеждна защитавътрешното съдържание от увреждане го прави важен компонент в такъв въпрос като структурата на клетката. Клетъчната мембрана на животинските организми понякога се оказва покрита с най-тънкия слой - гликокаликс, който включва протеини и полизахариди. Растителните клетки извън мембраната са защитени клетъчна стена, който изпълнява функциите на подкрепа и поддържане на форма. Основен компонент в състава му са фибри (целулоза) - полизахарид, който е неразтворим във вода.
По този начин външната клетъчна мембрана изпълнява функцията на възстановяване, защита и взаимодействие с други клетки.

Структурата на клетъчната мембрана
Дебелината на тази подвижна обвивка варира от шест до десет нанометра. Клетъчната мембрана на клетката има специален състав, чиято основа е липидният двоен слой. Хидрофобните опашки, които са инертни към водата, са разположени отвътре, докато хидрофилните глави, които взаимодействат с водата, са обърнати навън. Всеки липид е фосфолипид, който е резултат от взаимодействието на вещества като глицерол и сфингозин. Липидното скеле е плътно заобиколено от протеини, които са разположени в непрекъснат слой. Част от тях са потопени в липидния слой, останалите преминават през него. В резултат на това се образуват водопропускливи зони. Функциите, изпълнявани от тези протеини, са различни. Някои от тях са ензими, останалите са транспортни протеини, които пренасят различни вещества от външната среда в цитоплазмата и обратно.
Клетъчната мембрана е проникната и тясно свързана с интегрални протеини, докато връзката с периферните е по-слаба. Тези протеини изпълняват важна функция, която е да поддържат структурата на мембраната, да приемат и преобразуват сигнали от околната среда, да транспортират вещества и да катализират реакциите, протичащи върху мембраните.
Съединение
Основата на клетъчната мембрана е бимолекулен слой. Благодарение на своята непрекъснатост, клетката има бариерни и механични свойства. На различни етапитози двоен слой може да бъде нарушен в своите жизнени функции. В резултат на това се образуват структурни дефекти на хидрофилни пори. В този случай абсолютно всички функции на такъв компонент като клетъчната мембрана могат да се променят. В този случай ядрото може да страда от външни влияния.

Имоти
Клетъчната мембрана на клетката има интересни функции. Поради своята течливост тази обвивка не е твърда структура и по-голямата част от протеините и липидите, които съставляват нейния състав, се движат свободно в равнината на мембраната.
Като цяло клетъчната мембрана е асиметрична, така че съставът на протеиновия и липидния слой е различен. Плазмените мембрани в животинските клетки имат гликопротеинов слой от външната страна, който изпълнява рецепторни и сигнални функции, а също така играе важна роля в процеса на комбиниране на клетките в тъкан. Клетъчната мембрана е полярна навънзарядът е положителен, а отвътре е отрицателен. В допълнение към всичко по-горе, клетъчната мембрана има селективна проницателност.

Това означава, че освен вода, в клетката се допускат само определена група молекули и йони от разтворени вещества. Концентрацията на вещество като натрий в повечето клетки е много по-ниска, отколкото във външната среда. За калиевите йони е характерно различно съотношение: техният брой в клетката е много по-висок, отколкото в околната среда. В тази връзка натриевите йони са склонни да проникнат през клетъчната мембрана, а калиевите йони са склонни да се освобождават навън. При тези обстоятелства мембраната активира специална система, която изпълнява "изпомпваща" роля, изравнявайки концентрацията на веществата: натриевите йони се изпомпват към клетъчната повърхност, а калиевите йони се изпомпват навътре. Тази функция е включена в най-важните функции на клетъчната мембрана.

Тази тенденция на натриевите и калиеви йони да се движат навътре от повърхността играе голяма роля в транспортирането на захар и аминокиселини в клетката. В процеса на активно отстраняване на натриевите йони от клетката, мембраната създава условия за нови притоци на глюкоза и аминокиселини вътре. Напротив, в процеса на прехвърляне на калиеви йони в клетката, броят на "преносителите" на разпадните продукти от вътрешността на клетката към външната среда се попълва.
Как се подхранва клетката през клетъчната мембрана?
Много клетки приемат вещества чрез процеси като фагоцитоза и пиноцитоза. При първия вариант от гъвкава външна мембрана се създава малка вдлъбнатина, в която се намира уловената частица. След това диаметърът на вдлъбнатината става по-голям, докато заобиколената частица навлезе в клетъчната цитоплазма. Чрез фагоцитоза се хранят някои протозои, като амеба, както и кръвни клетки- левкоцити и фагоцити. По същия начин клетките абсорбират течност, която съдържа необходимите хранителни вещества. Това явление се нарича пиноцитоза.
Външната мембрана е тясно свързана с ендоплазмения ретикулум на клетката.
В много видове основни тъканни компоненти, издатини, гънки и микровили са разположени на повърхността на мембраната. Растителните клетки от външната страна на тази черупка са покрити с друга, дебела и ясно видима под микроскоп. Фибрите, от които са направени, помагат за поддържането на тъканите. растителен произход, например дърво. Животинските клетки също имат редица външни структури, които се намират върху клетъчната мембрана. Те имат изключително защитен характер, пример за това е хитинът, съдържащ се в покривните клетки на насекомите.
В допълнение към клетъчната мембрана има вътреклетъчна мембрана. Неговата функция е да разделя клетката на няколко специализирани затворени отделения – отделения или органели, където трябва да се поддържа определена среда.

По този начин е невъзможно да се надценява ролята на такъв компонент на основната единица на живия организъм като клетъчната мембрана. Структурата и функциите предполагат значително разширяване на общата клетъчна повърхност, подобряване на метаболитните процеси. Тази молекулярна структура се състои от протеини и липиди. Отделяйки клетката от външната среда, мембраната осигурява нейната цялост. С негова помощ междуклетъчните връзки се поддържат на достатъчно силно ниво, образувайки тъкани. В тази връзка можем да заключим, че една от най-важните роли в клетката играе клетъчната мембрана. Устройството и изпълняваните от него функции са коренно различни в различните клетки в зависимост от предназначението им. Чрез тези характеристики се постига разнообразие от физиологична активност на клетъчните мембрани и тяхната роля в съществуването на клетките и тъканите.