Органични киселини, липиди, витамини. Основи на липидната биохимия в човешкото тяло
Спиралните (организирани) участъци от нуклеинови киселини са оптично активни, въртят равнината на поляризирана светлина, абсорбират в ултравиолетовите лъчи при максимум 260 nm, но интензитетът на абсорбция от нуклеинови киселини е по-нисък, отколкото от смес от нуклеотиди - хипохромен ефект .
^ Лекция №3 (2 часа)
Тема: Биохимия на въглехидратите
Преди около сто години беше предложено въглехидратите да се наричат съединения, чийто състав съответства на формулата (CHOH) n, тоест въглеродни хидрати. Освен това n, тоест броят на въглеродните атоми, в зависимост от вида на въглехидратите, може да варира от три до няколко хиляди. Биологичните функции на въглехидратите са разнообразни и зависят от вида на въглехидратите или техните производни. Въглехидратите съставляват около 2% от масата на животинското тяло (в растенията въглехидратите съставляват до 80%). Най-голямо количество въглехидрати има в черния дроб, в съединителната тъкан. Основните функции на въглехидратите са:
Енергия (при разделянето на 1 g въглехидрати се образуват 4,1 - 4,2 kcal;
Пластмасови или структурни (конструктивни);
Защитни (в комбинация с протеини);
Регулаторни (също в комбинация с протеини);
Рецептор (също в комплекс с протеини) и др.
Всички въглехидрати са разделени на следните класове:
1. Монозахариди (мономерни единици), те понякога се наричат прости захари. Като цяло "захарид" - в превод от гръцки - "сладък". И наистина, много прости захари имат сладък вкус.
2. Олигозахариди (два или повече мономера, свързани в една верига).
3. Полизахариди - съединения с високо молекулно тегло, съдържащи от сто до няколко хиляди мономера.
Монозахариди
За първи път структурата на някои монозахариди е установена през втората половина на 19 век от Фишер. Всички прости монозахариди имат обща емпирична формула (CHOH) n, където n е цяло число от три до девет. Независимо от броя на въглеродните атоми, всички захариди могат да бъдат класифицирани в един от двата класа: алдози или кетози (окончанието -ose означава, че принадлежат към въглехидратите). Алдозите съдържат алдехидна функционална група, докато кетозите съдържат функционална група. Алдо- и кето-триозите, тетрозите, пентозите и др. се отличават по броя на въглеродните атоми. Разделянето на алдози и кетози може да се извърши и на друга основа: ако карбонилната група (= C =) е разположена в края на веригата, тогава това е алдоза, а ако карбонилната група е разположена някъде другаде, тогава това е кетоза или, съответно, алдехид или кетон.
^
Животинските тъкани съдържат следните монозахариди:
триози

В природата преобладават захари с D-конфигурация:
тетроси

Броят на изомерите за монозахаридите е 2 m, където m е броят на асиметричните въглеродни атоми. В триозите това е С 2, в тетрозите - С 2, С 3, в пентозите - С 2, С 3, С 4 и в хектозите - С 2, С 3, С 4, С 5 \u003d 16 изомери.
пентози

![]()
хексози


хептози

Всички изброени монозахари влизат в организма с храната или се образуват в метаболитни процеси. Те са безцветни, кристални твърди вещества, силно разтворими във вода. Монозахарта осигурява по-голямата част от енергията и по-голямата част от въглерода, необходими за синтеза на протеини, мазнини и други въглехидрати.
По правило монозахаридите с малък брой въглеродни атоми (триози, тетрози) имат структура под формата на прави вериги. А монозахаридите с брой въглеродни атоми 5 и повече имат затворена циклична структура; за да напишете тази структура, използвайте формулите на Haworth (1929)

Заради приликата с органичното вещество пиран  Глюкозата, записана с помощта на формулата на Heors, се нарича -D-глюкопироназа. Всичко, което се намира отдясно на въглеродните атоми при писане на формулата на Фишер, се намира отдолу при писане на формулата на Хехорс. и се определят от местоположението на групата - OH при първия въглероден атом (дясно или долу - ; ляво или горе - ). Последната CH 2 OH група (при последния С атом) винаги се записва най-отгоре.
Глюкозата, записана с помощта на формулата на Heors, се нарича -D-глюкопироназа. Всичко, което се намира отдясно на въглеродните атоми при писане на формулата на Фишер, се намира отдолу при писане на формулата на Хехорс. и се определят от местоположението на групата - OH при първия въглероден атом (дясно или долу - ; ляво или горе - ). Последната CH 2 OH група (при последния С атом) винаги се записва най-отгоре.
Петчленните пръстени се характеризират със сходство с химичното съединение фуран
 ,
следователно монозахаридите, записани в тази форма, се наричат фуранози:
,
следователно монозахаридите, записани в тази форма, се наричат фуранози:

В допълнение към монозахаридите, телесните тъкани съдържат и техните производни, които се образуват по време на реакциите на захари с помощта на реактивни групи:
OH, - C \u003d O, \u003d C \u003d O, т.е. алкохол, алдехид и кетон.
гликозиди
Монозахаридите са способни да образуват гликозидна връзка от типа C-O-R. Най-често гликозидната връзка се образува от алкохолна група (-OH) при първия въглероден атом, въпреки че други групи (-OH) могат да образуват гликозидна връзка при по-строги условия. Например:

Тоест всъщност това е реакцията на два алкохола, образуващи прост етер R–O–R. С помощта на гликозидна връзка монозахаридите могат да бъдат свързани помежду си:

Олигозахариди
Олигозахаридите включват захари, състоящи се от 2, 3, 4 или повече монозахариди. Лактозата се намира в тъканите (млечната захар – в кърмата е около 7%). Това е смесен олигозахарид (т.е. състои се от различни монозахаридни остатъци), по-точно дизахарид и се състои от глюкозни и галактозни остатъци, свързани с α-глюкозна връзка:

Смесените олигозахариди се намират в свободна форма (в кърмата) и свързани с протеини и липиди (в тъканите). Те включват глюкоза, галактоза, маноза, аминозахари, техните ацетилови производни, невраминова и сиалова киселина, L-фруктоза - това произвежда три-, тетра, - пентазахариди и др. Наличието на олигозахаридни групи в кръвта и тъканите определя груповата специфичност ( кръвни групи), те предизвикват специфични междуклетъчни взаимодействия. Малтоза, захароза и други олигозахариди се намират в растенията.
Бифидус факторът също принадлежи към смесените олигозахариди, включва галактоза, L-фукоза, N-ацетилглюкозамин, глюкоза и невраминова киселина. Този фактор е най-важният субстрат за ферментационната микрофлора.
полизахариди
Всички полизахариди могат да бъдат разделени на:
1. хомополизахариди (всички остатъци на съставните мономери са идентични). Единственият представител в животинските тъкани е гликогенът. Гликогенът служи като резервен енергиен материал за клетката. Състои се от глюкозни остатъци, свързани помежду си с -1,4-глюкозидна връзка. Всичко заедно се осъществява с помощта на -1,6-гликозидна връзка.

Разклоняването става чрез 8-10 глюкозни остатъка в главната верига. Разклоняването на гликогеновата верига увеличава нейната разтворимост и също така увеличава скоростта на синтез и разграждане на гликоген, тъй като зоната на контакт на гликогена с ензими (съответно гликоген синтаза и гликоген фосфорилаза) се увеличава.
Гликогенът присъства във всички клетки, но най-вече в черния дроб (5-10% от общата маса) и в мускулите (1-2%). Молекулното тегло на гликогена е до няколко милиона. Това е депо на глюкоза в организма.
2. Хетерополизахариди(съставени от различни мономери). Хетерополизахаридите почти винаги са в комплекс с едно или друго количество протеин (от 1 до 5%), като комплексите се наричат още протеогликани. Захаридната част на такъв комплекс се нарича мукополизахариди или гликозаминогликани. Има много протеогликани в съединителната тъкан (до 33%), особено в свободната съединителна тъкан (междуклетъчно вещество), сухожилията, връзките, кожата, роговицата, стъкловидното тяло, сърдечните клапи и др.
^ Смилане на въглехидрати
Това е процес на хидролитично разцепване на поли- и олигозахариди. Храносмилането започва в устата. Слюнката съдържа ензим, който разгражда нишестето (-амилаза), разгражда -1,4-гликозидните връзки в молекулата (но не крайните), като същевременно образува декстрини, малко количество малтоза и изомалтоза. В стомаха въглехидратите не се усвояват, т.к. pH е киселинно. Пълното храносмилане се извършва в тънките черва. Панкреасът произвежда , -амилази и ги освобождава в чревния лумен. Клетките на чревната лигавица също произвеждат редица ензими, които разграждат олигозахаридите. Една група ензими разцепва -гликозидните връзки (малтаза: -1,4-гликозидни връзки, изомалтаза--1,6; захараза - захароза). Втората група разцепва -гликозидните връзки (лактаза, -галактозидаза, хетерогалактозидаза - разцепва смесени олигозахариди). Под действието на тези ензими се получава разцепване до монозахариди. Храносмилането е по-често париетално (близо до клетъчната повърхност). Не се усвояват: фибри (без ензими), но са необходими за чревната подвижност, пектозани (полизахариди, състоящи се от пентози). По време на храносмилането се образува по-голямата част от глюкозата. Абсорбцията се извършва с помощта на специални носители, докато специфичността е характерна, изразходва се енергия (увеличава се консумацията на кислород, изразходва се АТФ). Глюкозата и галактозата могат да се абсорбират поради активен транспорт, поради концентрационния градиент на Na + йони, създадени от Na, K - ATPase. Това гарантира тяхното усвояване дори при ниски концентрации в червата. С кръвния поток през порталната вена глюкозата се доставя в черния дроб и частично с кръвния поток се доставя до всички органи и тъкани.
Глюкозата може да идва от стомашно-чревния тракт с кръвния поток, да се образува по време на разграждането на гликогена и може да се синтезира от невъглехидратни компоненти - глюконеогенеза.
гликолиза
Гликолизата обикновено се отнася до разграждането на въглехидратите. В зависимост от конкретния организъм и (или) условията на неговия растеж, гликолизата може да върви по анаеробния път (в този случай се нарича анаеробна гликолиза).
И може да следва аеробния път (съответно аеробна гликолиза). За много микроорганизми, растящи при анаеробни условия, гликолизата е основният катаболитен път за получаване на енергия от въглехидратни субстрати, което води до образуването на определени метаболитни крайни продукти, например етанол, лактат, глицерол. Този тип процес често се нарича ферментация. При животните анаеробната гликолиза (с образуването на лактат) осигурява енергия за свиване на скелетните мускули при условия на ограничено снабдяване с кислород. В някои случаи гликолизата (аеробна гликолиза) следва анаеробния път при аеробни условия, т.е. крайният продукт е лактат, въпреки че присъства кислород. Такива процеси протичат например в еритроцитите, ретината, чревната лигавица, в тъканите на плода (скоро след раждането). Гликолизата е единственият анаеробен път, който осигурява енергия. При аеробни условия (в присъствието на кислород), реакциите на гликолиза представляват първоначалната форма на разграждане на въглехидратите, допълнително свързана с цикъла на лимонената киселина. В този случай гликолизата спира на етапа на образуване на PVC. При аеробни условия гликолизата и цикълът на Кребс заедно водят до пълното окисление на хексозата до въглероден диоксид. Този процес е придружен от освобождаване на големи количества потенциално налична метаболитна енергия, главно под формата на високоенергийни продукти NADH и FADH 2, които, окислени по време на дихателната верига, осигуряват енергията, необходима за процеса на RP, което води до образуването на АТФ. Гликолизата е първата напълно дешифрирана последователност на био химична реакция, отнема почти 50 години (1890-1940). През това време са открити фосфоестери, АТФ, НАД, много фактори са изолирани и характеризирани и т.н. Гликолизата протича в цитозола.
Образованието е много важно финален етап NAD+, който се използва за окислително фосфорилиране на 3-фосфоглицералдехид.
Лактатът е крайният продукт на гликолизата. Тъй като лактатът (както и пируватът, между другото) преминава доста лесно през плазмената мембрана, той навлиза от мускулната клетка (или еритроцитите) в кръвта, с кръвния поток навлиза в черния дроб, където се окислява до пируват, който след това се превръща в глюкоза. Така се образува един вид цикъл. Тези трансформации съставляват цикъла на Кори:

Като цяло по време на гликолизата се образуват 4 молекули АТФ и се изразходват 2 АТФ. По този начин общият ефект от окисляването на една глюкозна молекула по време на гликолиза до лактат е 2 ATP. Използват се само около 3% от цялата възможна енергия, която би могла да се получи чрез окисляване на глюкозата, т.е. 27 kg глюкоза трябва да се разграждат на ден, за да се осигури енергия на тялото (ако енергията се получава само чрез гликолиза). Въпреки това, гликолизата се извършва в клетките. Необходим е в мускулите за бързо получаванеенергия.
^ Лекция №4 (2 часа)
Тема: Биохимия на липидите
Липидите са естествени неполярни съединения, които са напълно или почти напълно неразтворими във вода, но разтворими в неполярни разтворители, като хлороформ, въглероден дисулфид, етер и горещ етанол. Основната отличителна черта на тази група от доста хетерогенни вещества е тяхната неполярност, която в някои случаи се обяснява с наличието в тези съединения на една или повече мастни киселини, съдържащи дълги алифатни въглеводородни вериги. Съдържанието на липиди в организма е 10 - 20% от телесното тегло.
Липидите изпълняват следните биологични функции :
1. Те са компоненти на мембраните, които до голяма степен определят тяхната пропускливост.
2. Те служат като една от основните форми на съхранение на въглерод и енергия (1 g мазнини по време на окисление дава 9,3 kcal).
3. Могат да бъдат прекурсори на други важни съединения.
4. Те действат като защитни бариери, които предпазват от топлинни, електрически и физически влияния.
5. Те са част от защитни обвивки, които предпазват от инфекция и прекомерна загуба или натрупване на вода. Мазнините са източници на ендогенна вода.
6. В някои случаи това са витамини и хормони.
7. Участват в аддукцията на нервните импулси, тъй като са част от миелиновите обвивки.
Съществува класификация, базирана на съдържанието на един или друг алкохол в състава на мазнината. Съответно се изолират глицерин-съдържащи, сфингозин-съдържащи липиди и холестерол и неговите производни.
Класификация на липидите според тяхната структура:
1. Прости липиди.
2. Сложни липиди.
3. Липидни производни.
Простите мазнини включват само естери на мастни киселини и алкохоли. Съставът на комплексните липиди, освен мастни киселини и алкохоли, включва и други компоненти. Сред сложните липиди се разграничават фосфоацилглицероли, сфингомиелини, цереброзиди и ганглиозиди. Липидните производни включват всички съединения, които не могат да бъдат ясно класифицирани като прости или сложни липиди, като стероиди, каротеноиди и витамини от липиден характер. Много от тях се образуват от въглеводородни вериги, които са част от мастни киселини.Съставът на прости и сложни липиди задължително включва мастни киселини. Всъщност това са карбоксилни киселини с дълга алифатна верига. Естествените мастни киселини са много разнообразни.
За удобство се разграничават следните групи:
1. Повечето мастни киселини са монокарбоксилни киселини, съдържащи линейни въглеводородни вериги с четен брой атоми (обикновено C 12 - C 20). По-рядко срещани са ФК с по-къси вериги или с нечетен брой въглеродни атоми.
2. Често има киселини, съдържащи етиленови (ненаситени, двойни) връзки (обикновено C 18 - C 20 киселини). Ако има две или повече двойни връзки, те почти винаги са разделени от една метиленова група: - CH = CH-CH 2 -CH = CH -.
3. В ненаситените киселини с двойни връзки почти винаги има цис конфигурация.
Голям брой неполярни C-C и C-H връзки във въглеродната верига придават по същество неполярен характер на цялата молекула като цяло, въпреки че тя също има полярна група - COOH. Съвсем ясно е, че съединение, съдържащо един или повече FA остатъци, ще бъде неполярно, което е причината за неразтворимостта на липидите във вода и също така причинява сглобяването на липидите в биомембрани.
Обмисли различни видовелипиди:
^ 1. Прости липиди . Те се разделят на две групи: неутрални ацилглицероли и восъци.
Неутрални ацилглицероли.В зависимост от броя на FA остатъците, свързани с тривалентния алкохол глицерол, се разграничават моно-, ди-, триацил-глицероли. Триацилните производни са най-често срещаните в природата. Във всички случаи простият ацилглицерол не съдържа функционални йонни групи и принадлежи към неутралните липиди. Ацилните странични вериги обикновено са различни.

Восъци.Това са естери, но както алкохолът (до С 18), така и киселината, включени в състава им, имат дълги въглеводородни вериги. Всички восъци са напълно неразтворими във вода. Естествените восъци обикновено са крайните продукти на тези метаболитни пътища, чиято основна роля е да образуват защитни покрития. Птичите пера и животинските кожи имат восъчно покритие, което ги прави водоотблъскващи. Восъчното покритие на листата и плодовете на растенията намалява загубата на влага и намалява възможността от инфекция.
^ 2. Сложни липиди.
Има три основни класа сложни липиди:
1. Фосфоацилглицероли.
2. Сфингомиелини.
3. Гликолипиди.
Първите два класа се наричат фосфатиди или фосфолипиди поради наличието на фосфатни групи. Всички тези съединения се намират само в мембраните, като техни основни компоненти.
^ Фосфоацилглицероли (ПАГ). Това са най-често срещаните сложни липиди. Броят на различните ФК и тяхната относителна концентрация варира значително в зависимост от вида на клетките и тяхното физиологично състояние. Въпреки това разнообразие, те все още правят разлика между главни и второстепенни компоненти. Основен:фосфатилхолин (лецитин), фосфатидилетаноламин (цефалин), фосфатидилглицерин. Незначителен:фосфатидилсерин, дифосфатидилглицерол (кардиолипид), фосфатидилинозитол и фосфатидна киселина. Всички други FAGs се образуват от фосфатидна киселина.
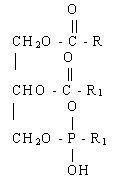
Фосфатна киселина
Различни FAG се различават един от друг по допълнителни групи, прикрепени чрез фосфоестерна връзка към фосфатидната киселина. Съставът на FA на различни FAG се различава дори в рамките на един и същи организъм.
^ Структура на някои ФА:


В тези структури ясно се вижда, че съединенията имат две различни области: неполярна хидрофобна „опашка“ и полярна хидрофилна „глава“. Такава двойствена природа се нарича алифатична (от гръцки - двустранно и чувство).
^ Сфингомиелини (фосфосвинголипиди). Те се състоят от една молекула мастни киселини, холин, фосфорна киселина и суингозинов алкохол. Сфингозините са семейство аминоалкохоли с дълга ненаситена верига, които се различават по дължина на веригата. FA се прикрепя към аминогрупата на сфингозин чрез амидна връзка и се образува керамид. Добавянето на фосфорилхолинов остатък завършва образуването на суингомиелин:

^ Гликолипиди (гликосфинголипиди). Те също са на базата на керамид. Има цереброзиди и ганглиозиди. Друга последователна липидна биосинтетична реакция е ензимното превръщане на керамидните прекурсори в цереброзиди и ганглиозиди чрез свързване на въглехидратна група към керамида. Цереброзидите и ганглиозидите се намират главно (макар и не изключително) в клетъчните мембрани на нервната и мозъчната тъкан и също проявяват амфипатични свойства. Протозои цереброзидисъдържат монозахаридна група, свързана чрез гликозидна връзка към крайната -ОН група на церамида. По правило това е глюкоза или галактоза.

^ 3. Липидни производни.
Това е много разнородна група от съединения. Тяхното сходство е ограничено само от слабата разтворимост във вода. Най-важните членове на тази група са стероиди, каротеноиди и сродни витамини от липиден характер.
Стероиди.Те се намират във всички организми, където изпълняват различни функции. При хората те играят ролята на полови хормони, емулгиращи агенти при смилането на липидите и участват в транспорта на липидите през мембраните. В кръвната плазма те действат като противовъзпалителни средства и като регулатори на някои метаболитни процеси. Всички стероиди имат подобна структура, която се основава на перхидроциклопентанофенантрен. Разнообразието от стероидни структури се дължи на различни степениненаситеност и наличието на няколко групи в различни позиции на циклите.

Наличието на въглеводородна странична верига (C 8 - C 10) в 17-та позиция и хидроксилна група в позиция 3 е характерно за голяма група стероиди, наречени стероли. Най-важният и често срещан от тях е холестеролът. Той влиза като структурен компонент в клетъчните мембрани, но концентрацията му в мембраните варира (до 40% от общото съдържание на мембранни липиди). Холестеролът (поради съдържанието на консолидирана структура, която е по-малко гъвкава от разширената въглеводородна верига) придава на мембраната по-голяма твърдост (якост). Холестеролът също е основният метаболитен прекурсор на други важни стероиди, включително жлъчни киселини и полови хормони. В някои тъкани холестеролът е предшественик на витамин D. Половите стероидни хормони: мъжки - тестостерон (причинява всички полови белези) и женски: естрадиол и прогестерон (основно отговорни за регулирането на менструалния цикъл).

За първи път е изолиран от камъни в жлъчката, откъдето идва и името (chole - жлъчка, steros - твърд).
Каротеноиди.Каротеноидите се разделят на две групи: каротини и ксантофили. И двете групи включват водонеразтворими пигменти, които са широко разпространени в природата, повечето от тях се намират в растения и водорасли. Каротините са чисто въглеводородни съединения, докато ксантофилите съдържат кислородни групи. Каротините са по-широко разпространени в природата. Най-често срещаният е бета-каротинът. Това е C 40 въглеводород със силно разклонена структура и ненаситени връзки, в двата края на въглеводородната верига има идентични заместени пръстени. Всички други каротеноиди по същество могат да се разглеждат като варианти на тази структура. По време на ензимното разцепване на бета-каротин се образуват две молекули на витамин А по симетричен начин.
^ Липидни витамини. Витамин D.Необходим за нормалния метаболизъм на калция и фосфора, необходими за растежа на здрави кости и зъби. Недостигът му причинява рахит – заболяване, при което костите стават меки и пластични, което води до тяхната деформация. Витамин D се образува от прекурсор на стерол с UV лъчение. Един важен прекурсор е 7-дихидрохолестеролът, който сам по себе си се произвежда ензимно от холестерола. От него се получава витамин D3. Има няколко форми на витамин D. Форма D 3 (холикалцеферол) присъства в млякото, маслото, черния дроб на рибите, които са основният екзогенен източник на този витамин. Витамин D3 може да се синтезира в кожата, при условие че се получава нормална порция слънчева светлина.
Витамин Е.Основната структура на витамин Е (и има няколко форми) се нарича токоферол. Най-активната форма на витамин Е е -токоферол. При плъховете витамин Е е необходим за репродукцията (това все още не е доказано за хората). При някои животни липсата на витамин Е води до мускулна дистрофия. Витамин Е обикновено се добавя по време на обработката на храната, тъй като предотвратява развалянето поради своите антиоксидантни свойства. хранителни продуктипоради окисляването им. В живите клетки витамин Е също действа като антиоксидант, заедно с витамин С и глутатион.
Витамин К.Недостигът му забавя съсирването на кръвта. Структурата му се основава на бициклична нафтохинонова система, към която е прикрепена дълга въглеводородна верига. То може да дойде от храна (пресни зеленчуци, сирене) или може да бъде стимулирано от бактерии в стомашно-чревния тракт. Неговият коагулационен ефект се определя от участието в синтеза на протромбин, който след това преминава в тромбин.
Биомембрани
Биомембраните обхващат клетката и много органели в клетката. Много е активно биологични системи, отговорен за такива процеси като селективен транспорт на вещества в и извън клетката и субклетъчните отделения (формации), свързването на хормони и други регулаторни молекули, реакции, катализирани от ензими, предаване на електрически импулси и дори синтеза на АТФ . Мембраните са различни и различни видовемембраните имат различни видове функционална дейност. Например, само вътрешната митохондриална мембрана е отговорна за биосинтезата на АТФ.
^ Химичен състав. Мембраните са съставени от липидни и протеинови молекули. Относителното им количество се различава значително за различните мембрани, като варира от 20% протеин + 80% липиди до 75% протеин + 25% липиди. Въглехидратите под формата на гликопротеини и гликолипиди съставляват 0,5 - 10% от мембранното вещество.
^ Липидно храносмилане
Наскоро имаше съобщения за съществуването на липаза, секретирана от дорзалната повърхност на езика, която участва в смилането на диетичните триацилглицероли. Липазата също се секретира в стомаха, но поради ниското pH, тази липаза е от съществено значение физиологично значениене притежава. В същото време липазата на езика може да запази действието си от два до четири часа в хранителния болус. Липазата на езика е най-активна по отношение на късоверижните мастни киселини и действа предимно на 3-та позиция. Особено добър субстрат за този ензим е млечната мазнина (съдържа главно късоверижни и средноверижни мастни киселини, естрифицирани на 3-та позиция). Основното храносмилане на липидите става в тънките черва. Всъщност смилането на мазнините се свежда до хидролитично разцепване. Това изисква следните условия:
1. Наличие на подходящи ензими (липази);
2. Оптимално pH (5 - 7);
3. Мазнините трябва да бъдат емулгирани (т.е. натрошени), тъй като във водната среда те са под формата на големи капки, които са недостъпни за действието на ензими, които, тъй като са водоразтворими, не могат да проникнат в капката. Ако тази капка се смачка (емулгира), тогава общата повърхност >> и съответно >> действието на ензима. Липазите разцепват мастните киселини от триацилглицеролите в позиции 1 и 3. Най-трудно е деколтето на 2-ра позиция. Този процес се катализира от ензима фосфолипаза А 2 . По този начин, при пълно смилане, триацилглицеролите се разграждат до глицерол и мастни киселини. Но, както вече споменахме, за пълното разграждане на мазнините в тънките черва, мазнините трябва да бъдат емулгирани. По принцип емулгирането се свежда до:
1. Намалено повърхностно напрежение;
2. Раздробяване;
3. Стабилизиране на емулсията (покриване на малки капчици мазнина с протеинова обвивка). Важна роля в емулгирането на мазнините играят жлъчните киселини, които се образуват от холестерола.
^ Лекция №5 (2 часа)
Тема: Структурна организация на клетките и органните системи, свързани с храненето
клетъчна структура
клеткае основната елементарна единица на живия обект. Срок жив обектобикновено включва всички обекти, способни на метаболизъмИ размножаванекато себе си.
За поддържане на тези две основни функции в процеса на еволюцията в клетката са възникнали определени структури, т.нар клетъчни органели. Те осигуряват координирано и контролирано протичане на основните реакционни процеси, необходими за постоянното проявление жизнени функции.
Следните клетъчни органели са важни за съществуването на живия организъм: ядро, митохондрии, ендоплазмен ретикулум, рибозоми, лизозоми и микротелца. Клетъчни мембрани, не само отделят жив организъм (клетка) от заобикаляща среда, но участват в образуването на определени дялове на клетката (функционални деления). Те служат като структурен елемент на всички клетъчни органели и участват във функционирането на повечето от тях. Масата на мембраните може да достигне 80% от масата на клетката. Неструктурирана, колоидна маса, която изпълва вътреклетъчното пространство, се нарича цитозол.
Понякога клетката съдържа морфологично различни гранули, съдържащи продукти от клетъчната активност, или продукти за съхранение (гликоген, мастни капки) или продукти, които трябва да бъдат транспортирани извън клетката (проензими).
Независимо живите клетки (едноклетъчни организми) обикновено съдържат всички изброени по-горе структури и освен това имат клетъчна стена и в някои случаи съкратителен апарат (реснички и флагели).
При многоклетъчните организми има разлика във функциите, основана на диференциацията на структурите. По този начин в диференцираните клетки на висшите организми има разлики в броя на клетъчните органели (понякога разлики в тяхната фина структура, например в броя на кристите в митохондриите на клетките на черния дроб и сърдечния мускул), както и в техните различно разпределение в клетката (например натрупване на митохондрии в клетката, където протичат ендергонични процеси).
Външната част на плазмената мембрана включва сложни химични структури (протеини, гликопротеини) и се нарича гликокаликс. Тези структури служат за взаимно разпознаване от клетки на даден вид и за разпознаване на клетки от други видове. Ако тези структури са част от специално диференцирани клетки на висши организми, те служат като антигени и предизвикват образуването на различни специфични антитела.
Дори в органите клетките обикновено не са в близък контакт. Това се дължи на наличието на отрицателни заряди на повърхността на клетката, които се отблъскват взаимно. В резултат на това се образуват тесни пространства между клетките, чийто сбор в целия орган или организъм се означава като междуклетъчно пространство. По същия начин сборът от всички компоненти в клетката (напр. ядро, митохондрии и т.н.) се нарича вътреклетъчно пространство.
От функционална гледна точка не може да има живи организми (или дори клетки или техните органели) с неограничено време на съществуване.
Започвайки от процеса на делене, всички клетки преминават през т.нар жизнен цикъл, в края на който или настъпва делене с появата на нова клетка, или настъпва смърт. Продължителността на този цикъл е видоспецифична и според съвременните представи варира от няколко часа до десетки години. По време на жизнен цикълклетката преминава през определени фази, които продължават различно време в зависимост от вида на клетката и се характеризират със строго специфични метаболитни процеси. Тези фази са означени като G1, S, G2 и т.н. Клетките, които не се делят по-нататък и умират след определено време (например клетки от сивото вещество на мозъка), са постоянно във фаза G 1. Към тази група клетки принадлежат така наречените диференцирани клетки.
Фактът, че някои от ДНК молекулите в ядрото са свързани с хистони, преди това се считаше за молекулярна основа. диференциация. Тази гледна точка беше отхвърлена, след като беше показано, че хистоните играят решаваща роля в образуването на суперспирала, във фибрилите на която те са част от дезоксирибонуклеопротеините и се освобождават само в процеса на репликация на ДНК.
В съответствие със съвременните концепции процесът на клетъчна диференциация е непрекъснат, той е напълно необратим поради функционалното блокиране на гените. Предполага се, че този процес трябва да се контролира от продуктите на гените, отговорни за структурата и времето на синтеза.
^ Описание на типична клетка
Ядро, заобиколен от двойна мембрана с пори, е локализиран в средата на клетката. Вътре в ядрата се виждат тъмни нуклеоли. Външната ядрена мембрана е част от ендоплазмения ретикулум, свързан с Комплекс Голджи. Рибозомите са разположени на повърхността ендоплазмения ретикулум. Има овални структури, заобиколени от двойна мембрана, чиято вътрешна част образува кристи митохондриите. Лизозомизаобиколен от един слой мембрана. Те съдържат хидролитични ензими, повечето от които са неактивни като проензими. В едноклетъчните организми те са отговорни за смилането на веществата, които влизат в клетката. Във висшите организми лизозомите участват в разграждането на клетки, които са престанали да изпълняват своите функции. микрозоми (пероксизоми)) са по-малки от лизозомите. Те съдържат оксидази, които катализират окисляването на съединения, които са чужди за клетката и следователно трябва да бъдат отстранени от нея (напр. лекарства, ароматни съединения и др.). Клетка заобиколена плазмената мембрана, който е конструиран по такъв начин, че на определени места става възможно директно прехвърляне на съединения от извънклетъчното пространство към ядрото. Пространството между органелите, изпълнено с колоидна суспензия, богата на протеини (ензими), се нарича цитозол.

плазмената мембрана
Плазмената мембрана се образува от протеини (периферни и интегрални), вградени в липиден двоен слой. ^ интегрални протеини са гликопротеинов характер. Тяхната N-терминална част е част от вътрешния фосфолипиден слой, в който прониква част от пептидната верига, богата на неполярни аминокиселини (в спирална конформация), а страничните им вериги влизат в множество хидрофобни контакти с алифатни фосфолипидни вериги. Олигозахаридните вериги, обикновено съдържащи GlcNAc, Man, Cal, Fuc и N-ацетилневраминова киселина, могат да бъдат свързани към интегрална протеинова пептидна верига на външната повърхност на плазмената мембрана. N-ацетилневраминовата киселина обикновено стои в края на олигозахаридната верига и причинява нейния отрицателен заряд. Олигозахаридите придават специални свойства на клетъчната повърхност, позволявайки й да разпознава клетки от същия орган или клетки от друг тип (антигенност, контактно инхибиране). Олигозахаридите образуват слой върху клетъчната повърхност, наречен гликокаликс. Структурите, локализирани на клетъчната повърхност, предотвратяват близкия контакт между клетките. Това води до факта, че между клетките се появява повече или по-малко тясно пространство, пълно с течност. Общото наименование на такива места в даден орган или организъм е междуклетъчното пространство. Сумата от всички обеми вътре в клетките се нарича вътреклетъчно пространство.

Митохондриите
Това е мястото на образуване на АТФ. Енергията, необходима за неговия синтез, се появява в резултат на постепенното окисление на водород-съдържащите субстрати (захари, липиди, аминокиселини) в дихателната верига под действието на кислород. Декарбоксилирането в цикъла на лимонената киселина води до образуването на CO 2, резултатът от окислението е образуването на H 2 O. Ензимите, които осигуряват пренос на електрони, са част от вътрешната мембрана на митохондриите. Кислородът навлиза в митохондриите чрез дифузия. Продуктът на митохондриалната активност (АТФ) се пренася чрез процеси на транслокация от мястото на неговото образуване до екстрамитохондриалното пространство, където се използва. За да се осигури бърз трансфер на АТФ, митохондриите се локализират в близост до структури, където протичат енергоемки процеси (например близо до елементи, участващи в процеса на свиване).

Интерфазно ядро и хромозоми
Интерфазното ядро е изпълнено с вещество, наречено хроматин. Заедно с ДНК в хроматина присъстват два вида протеини: основните протеини са хистони и нехистонови протеини (които по правило са киселинни). Хроматинът се състои от повтарящи се структурни елементи - нуклеозоми (или v-тела). Ядрото на нуклеозомата се формира от четири типа хистони, давайки октамер, съдържащ две молекули от всеки от хистоните H3, H4, H2A и H2B. Хистоновите молекули са свързани една с друга чрез хидрофобни взаимодействия и техните N-терминални последователности (главно положително заредени) са разположени на повърхността на октамера. Това от своя страна осигурява взаимодействието на октамера с двойната спирала на ДНК. Така свързаният ДНК фрагмент съдържа в зависимост от биологичния обект 154-241 базови двойки. Разстоянието между две уклеозоми е в рамките на 9–14 nm, а ДНК, разположена в този регион, е свързана с хистон H1. Генетичната информация, съдържаща се в даден участък от ДНК, става достъпна само след модификация на хистоновата молекула (например след нейното фосфорилиране).
След това ДНК молекулите, свързани с нуклеозомите, се навиват в суперспирала. По този начин дори отдалечени ДНК сегменти могат да бъдат в непосредствена близост, образувайки, например, прекъснати гени. Преди клетъчното делене хроматинът е в тази най-кондензирана форма и образува хромозоми. Предполага се, че функцията на нехистоновите протеини се проявява в процеса на транскрипция.
Количеството ДНК в клетка е постоянно (6 pg в клетка на бозайник). Тази стойност съответства на 5,5 х 109 нуклеотидни двойки. Молекулното тегло на ДНК е 10 10 -10 11. Дължината на една напълно разтегната молекула би трябвало да бъде няколко сантиметра. В хромозомите молекулите на ДНК са в силно кондензирана форма (44:1), а in vivo 7 10 -6 g ДНК съответстват на 1 микрон.

Рибозоми, полизоми
Рибозомите и полизомите са сферични и се намират в цитоплазмата или свободни, или свързани с мембраните на ендоплазмения ретикулум. Рибозомите са изградени от две субединици. По време на протеиновия синтез иРНК се свързва с малка субединица. Няколко рибозоми (не по-малко от 4 и не повече от 100) могат да се прикрепят към една иРНК молекула. Този комплекс се нарича полизома (полирибозома). Рибозомите могат да се разпаднат на субединици; този процес зависи от концентрацията на магнезиевите йони. Всяка субединица е изградена от рРНК молекула и специфичен набор от протеини. Броят на рибозомите в бактериална клетка достига 10 4, в животинска клетка е около 10 5.

Ендоплазмения ретикулум
Ендоплазменият ретикулум е мембранна структура, разположена в цитоплазмата, близо до ядрото. В електронен микроскоп се виждат тръби, наречени цистерни, на чиято външна повърхност могат да бъдат разположени рибозоми. Следователно тези структури се наричат гранулиран или груб ендоплазмен ретикулум (SER), за разлика от гладкия ендоплазмен ретикулум (SER), който няма свързани рибозоми. Рибозомите са прикрепени към страната на цитоплазмата, където се извършва протеиновият синтез. След завършване на протеиновия синтез, получените пептидни вериги преминават през мембраната в цистерната и се прехвърлят на определени места в клетката или в комплекса на Голджи. Според текущата гледна точка тези протеини, които трябва да бъдат отстранени от клетката и, вероятно, протеиновите компоненти на мембраните, се синтезират в грубия ендоплазмен ретикулум, а протеините, които се използват от клетката, се синтезират върху свободни рибозоми.

Комплекс Голджи
Комплексът на Голджи се образува от паралелни тубулни мембранни системи, които са директно свързани с грапавия ендоплазмен ретикулум. специфични ензими, гликозилтрансферази, катализиращи свързването на монозахариди с протеини с помощта на гликозидни връзки (чрез Ser или Thr OH групите, по-рядко чрез Asn амидната група), са част от мембраните на апарата на Голджи. Монозахаридите обикновено участват в този процес под формата на производни с UDP или CMP. След като прикрепи въглехидратната част, протеиновата молекула може да напусне клетката поради процеса екзоцитоза. Но преди това да се случи, протеинът трябва да остане в клетката за определено време. Поради това съдържанието на вакуолите на комплекса на Голджи постепенно се концентрира (водата се елиминира) и протеиновите продукти (дори в кристална форма) се отлагат под формата на гранули.

Моторният апарат на клетката
Както едноклетъчните, така и многоклетъчните организми могат да бъдат снабдени с реснички и флагели, разположени на повърхността, осигуряващи движение на организма или неговата среда. Тези структури са изградени от фибриларни протеини (съдържащи глобуларни субединици), които под въздействието на макроергични съединения могат да променят своята пространствена структура (конформация). Тази промяна в конформацията на протеиновата молекула е координирана и регулирана. Ултраструктурата на ресничките и флагелите е еднаква при всички еукариоти. Те съдържат девет периферни двойки влакна и една централна двойка. Всяка нишка е изградена от дванадесет или повече протофибрили. Всяка протофибрила е образувана от глобуларни субединици с диаметър около 4,5 nm. Протеините, наречени динеини, бяха изолирани от външните нишки. Те проявяват АТФазна активност в присъствието на АТФ и Mg 2+ и промяната в тяхната конформация изглежда важна за движението. Цитоплазмените микротубули имат подобна структура.
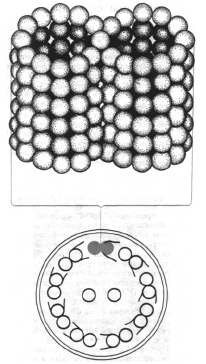
Клетъчен растеж и делене
При деленето на майчината клетка се получават две дъщерни клетки. През последващия период клетките растат и се подготвят за по-нататъшно делене. Интервалът между две митози ( клетъчен цикъл ) във фазата на експоненциален растеж е 10 минути за бактериална клетка и 24 часа за животинска клетка. През това време клетката преминава през няколко фази на растеж. IN постмитотична фаза G 1 клетката синтезира РНК молекули и протеини (ДНК не се синтезира). Продължителността на тази фаза е 30-40% от времето на целия цикъл. Клетките, които не се делят по-нататък (като мускулни и глиални клетки), са постоянно във фаза G 1. Във фазата на синтез (S) настъпва пълно дублиране на ДНК. В по-малка степен има синтез на РНК и протеини. Тази фаза отнема 30% от времето на цикъла. Подготовката за митоза се извършва в фаза G 2 . Подробностите за този метаболитен процес засега не са известни, но е ясно, че енергията, необходима за митозата, се съхранява по това време. РНК и протеините продължават да се синтезират и през това време. Тази фаза продължава 10-20% от времето на цикъла. Митозазаема 5-10% от цикъла и през това време метаболитните процеси не се случват.
В HeLa клетките общата продължителност на цикъла е 23 часа, G 1 =12 часа, S=5 часа, G 2 =5 часа, митоза =1 час.

БЕЛОРУСКИ ДЪРЖАВЕН УНИВЕРСИТЕТ ПО ИНФОРМАТИКА И РАДИОЕЛЕКТРОНИКА
Катедра ЕТТ
"Основи на липидната биохимия в човешкото тяло"
МИНСК, 2008 г
Липиди е общо наименование за всички известни мазнини и мастноподобни вещества с различна структура, но общи имоти(неразтворимост във вода, екстракция с неорганични разтворители). Липид (гръцка мазнина).
В човешкото тяло 10-20% мазнини от телесното тегло. Липидите са:
Протоплазмени - са част от всички клетъчни структури, органи и тъкани и практически остават на едно и също ниво през целия живот. Те съставляват 25% от всички телесни мазнини.
Резервните липиди се съхраняват в тялото и тяхното количество варира в зависимост от възрастта, пола, хранителните условия и дейностите.
Функции на липидите в организма:
1 - пластична функция: те участват в изграждането на клетъчните мембрани на всички органи и тъкани и образуването на много биологично важни съединения (хормони, мастноразтворими витамини).
2 - енергийна функция: липидите осигуряват 25-30% от енергийните нужди на тялото. Разграждането на 1 g мазнини е 9,3 kcal.
3 - мазнините са резервни хранителни вещества, тяхното депо е подкожна тъкан, периренална капсула.
4 – защитна функциялипиди: те участват в терморегулацията, предпазват кожата от изсушаване, органите - от сътресения.
5- действат като защитни мембрани, които предпазват от инфекция или прекомерна загуба или натрупване на вода.
6 - осигуряват усвояването на мастноразтворимите витамини
Класификация на липидите:
1 - прости или неутрални мазнини (естери на мастни киселини и алкохоли) Неутралните мазнини са в тялото или под формата на протоплазмена мазнина, която е структурен компонент на клетките, или под формата на резервна, резервна мазнина.
2 - сложни мазнини, представляват естери на тривалентния алкохол глицерол, високомолекулни мастни киселини и други компоненти. Сред сложните мазнини са: фосфолипиди, гликолипиди, сфигномиелини. Сфинголипидите се намират в мембраните на животински и растителни клетки.
3 - производни на липидите. Те включват всички съединения, които не могат да бъдат ясно класифицирани като прости или сложни липиди, като стероиди, каротеноиди и витамини от липиден характер.
4-восъци - например ланолин, смес от холестеролови естери.
Восъците са естери, образувани от наситени и ненаситени мастни киселини и алкохоли.
В неутралните мазнини се намират:
Мастна киселина.
Мастните киселини получават името си от начина, по който са изолирани от мазнините. Това са карбоксилни киселини с дълга алифатна верига.
Естествените мастни киселини са много разнообразни. Повечето мастни киселини са монокарбоксилни киселини, съдържащи линейни въглехидратни вериги с четен брой атоми. Съдържанието на ненаситени мастни киселини е по-високо от това на наситени. Ненаситените мастни киселини имат по-ниска точка на топене.
свойства на мастните киселини.
Наситените и ненаситените мастни киселини се различават значително по своята структурна конфигурация. В наситените мастни киселини въглеводородната опашка може по принцип да приеме различни конформации поради пълната свобода на въртене около крайната единична връзка.
При ненаситените киселини се наблюдава различна картина: невъзможността за въртене около двойната връзка осигурява твърд завой във въглеводородната верига.
Естествените мастни киселини, наситени и ненаситени, не абсорбират светлина нито във видимата, нито в UV област. Спектрофотометрично се определя само след изомеризация (230-260 nm). Ненаситените се определят чрез количествено титруване. Анализът на сложни смеси от мастни киселини се извършва чрез газово-течна хроматография.
Наситени - палмитинова, стеаринова, липоцеринова киселини
Ненаситени: арахидонова, олеинова, линолова, линоленова.
Растителните мазнини се състоят главно от ненаситени мастни киселини.
Липидите са задължителни интегрална частбалансирана човешка диета. Съотношението на протеини, липиди и въглехидрати трябва да бъде 1:1:4.
Стойността на мазнините е много разнообразна. Високото им съдържание на калории им придава специална стойност. Мазнините са разтворители на витамини А, D, Е и др. С мазнините в организма се въвеждат някои ненаситени киселини, които се класифицират като есенциални мастни киселини (линолова, линоленова, арахидонова), които не се синтезират при хора и животни. С мазнините в тялото влиза комплекс от биологично активни вещества: фосфолипиди, стероли.
Триацилглицероли – тяхната основна функция е съхранението на липиди. Те се намират в цитозола под формата на фини емулгирани маслени капчици.
Комплексни мазнини:
Фосфолипидите - основните компоненти на клетъчните мембрани и субклетъчните органели, изграждат по-голямата част от тъканите на мозъка, нервите, черния дроб, сърцето, участват в биосинтезата на протеини, активирането на протромбина, транспорта на липиди и мастноразтворими витамини в кръвта и лимфата. Състои се от глицерол и две молекули мастни киселини, едната от които е наситена. а другата е ненаситена + азотна основа.
Липопротеини.
Полярните липиди се свързват с определени специфични протеини, за да образуват липопротеини, от които транспортните липопротеини са най-известни и присъстват в кръвната плазма на бозайниците.
В такива сложни липиди взаимодействията между липидите и протеиновите компоненти се осъществяват без участието на ковалентни връзки.
Липопротеините обикновено съдържат както полярни, така и неутрални липиди, както и холестерол и неговите естери. Те служат като формата, в която липидите се транспортират от тънките черва до черния дроб и от черния дроб до мастната тъкан и други тъкани.
В кръвната плазма са открити няколко класа липопротеини, като тяхната класификация се основава на разликите в тяхната плътност. Липопротеини с различни съотношения на липиди и протеини могат да бъдат разделени в ултрацентрофуга.
Най-леките липопротеини са хиломикроните: големи структури, съдържащи около 80% триацилглицероли, 7% фосфоглицериди, 8% холестерол и неговите естери и 2% протеин.
Плазмените бета-липопротеини съдържат 80-90% липиди, а алфа-липопротеините - 40-70%.
Точната структура на липопротеините все още не е известна, но има причини да се смята, че протеиновата верига е разположена на външната повърхност, където образува тънка хидрофилна обвивка около мицеларната липидна структура. Съхранява се в мазнини или триглицериди повечето отенергия, освободена в резултат на химични реакции.
Наред с неполярните липиди има полярни липиди. Те са основните компоненти на клетъчните мембрани. Множество ензими и транспортни системи са локализирани в мембраните. Много свойства на клетъчните мембрани се дължат на наличието в тях на полярни липиди.
Мембранни липиди:
Мембранните липиди, заедно с въглеводородните вериги, съдържат една или повече силно полярни „глави“. Фосфолипидите присъстват в малки количества в мембраните. Техният основен компонент - фосфоглицеридите - съдържат 2 мастни киселинни остатъка, които естерифицират първата и втората хидроксилна група на глицерола. Третата хидроксилна група образува естерна връзка с фосфорната киселина. Хидролизира при нагряване с киселини и основи, както и ензимно - под действието на фосфолипази.
Сфинголипидите са вторият клас мембранни липиди, те имат полярна глава и две неполярни опашки, но не съдържат глицерол.
Те са разделени на 3 подкласа: сфингомиелини, цереброзиди и ганглеозиди.
Сфингомиелините се намират в миелиновите обвивки на някои видове нервни клетки. Церброзиди – в мембраните на мозъчните клетки. Ганглеозидите са важни компоненти на специфични рецепторни места, разположени на повърхността на клетъчните мембрани. Те се намират в онези специфични области на нервните окончания, където се осъществява свързването на невротрансмитерни молекули в процеса на химическо предаване на импулс от една нервна клетка към друга.
Външен или плазмени мембраниса изследвани много клетки, както и мембраните на редица вътреклетъчни органели, като митохондрии и хлоропласти. Всички мембрани съдържат полярни липиди.
Липидната част на мембраната е смес от полярни липиди. Естествените мембрани се характеризират с малка дебелина (6-9nm) и еластичност. Водата лесно преминава през мембраните, но те са практически непропускливи за замърсени йони като натрий, хлор или водород и за полярни, но незамърсени захарни молекули. Полярните молекули проникват с помощта на специфични носители на транспортната система.
Фосфоглицеридите, сфинголипидите, гликолипидите и восъците често се наричат осапуняеми липиди, тъй като при нагряването им (в резултат на елиминирането на мастни киселини) се образуват сапуни. Клетките също така съдържат по-малко неосапуняеми липиди, те не се хидролизират с освобождаване на мастни киселини.
Има 2 вида неосапуняеми липиди:
Стероиди и терпени
Стероиди - жлъчни киселини, полови хормони, надбъбречни хормони.
Стероидите са широко разпространени в природата. Тези съединения включват множество вещества от хормонален характер, както и холестерол, жлъчни киселини и др.
Стероли – Холестерол Холестеролът играе ролята на междинен продукт в синтеза на много други съединения. Холестеролът е богат на плазмените мембрани на много животински клетки; той се намира в много по-малки количества в мембраните на митохондриите и в ендоплазмения ретикулум.
Растенията съдържат фитостероли.
Терпени - намират се в растенията, много от тях придават на растенията характерния аромат и служат като основни компоненти на "ароматните масла".
мастноразтворими витамини.
Витамините са жизненоважни вещества, присъстващи в тялото в следи и необходими за нормалните клетъчни функции. Мастноразтворимите витамини включват витамини А, Е, К, D, молекулярната основа на тяхното действие не е точно известна.
Витамините се класифицират като липиди, защото са неразтворими във вода и могат да се екстрахират с органични разтворители. Мастноразтворимите витамини имат изопреноидна структура (A, E, K), витамин D е стероидно производно, въпреки че стероидите също идват от изопреноидни прекурсори. Витамин А се намира само в животинските тъкани. Съществува в 2 химични форми А-1 и А-2 (витамири) - ретинол 1 и ретинол 2 и представлява алкохол, съдържащ ацикличен пръстен, към който е прикрепена странична верига, състояща се от 2 изопренови единици.
Растенията съдържат каротеноидни пигменти. Алфа-, бета- и гама-каротините, при окислителното им разграждане в животинските тъкани се образува витамин А. Липсата на витамин А води до нарушен растеж и развитие на „нощна слепота“, нарушава се нормалната функция на пръчките на ретината.
Витамин Е е представен от цяла група витамини, присъстващи в растителните масла и наречени токофероли. Тези съединения имат ароматна пръстенна система, съдържаща хидроксил, и изопреноидна странична верига. Недостигът води до атрофия и мускулна слабост, стерилитет. Смята се, че тези вещества предотвратяват разрушителното действие на молекулярния кислород, понякога се наричат антиоксиданти.
Витамин К - К 1 и К 2, нафтохинони с дълги странични изопреноидни вериги с различна дължина. Недостигът се проявява в нарушение на процеса на съсирване на кръвта поради загубата на способността на организма да синтезира протромбин.
Витамин D е производно на стероиди. Най-важният е витамин D 2 - калциферол и D 3. Липсата на витамин D води до нарушения на метаболизма на калция и фосфора, което се проявява в промени в структурата на костите и зъбите. Витамин D насърчава абсорбцията на Ca йони в тънките черва чрез стимулиране на синтеза на протеин, участващ в транспорта на тези йони.
Простагландини.
Тези вещества се намират в почти всички органи и тъкани на хора и животни, тяхната висока активност, широк спектър на действие е сравнима с ефекта на хормоните.
Простагландините са циклични полиненаситени мастни киселини с относително молекулно тегло 300-400. Те съдържат само въглерод, кислород и водород.
Биосинтезата на простагландини се осъществява в микрозомите на клетката. Ненаситените мастни киселини са предшественици на простагландините. Те се синтезират при физиологична нужда. Тяхната роля е най-активните хранителни вещества. Те намаляват стомашната секреция, засягат гладката мускулатура, сърдечно-съдовата система.
Те се определят чрез абсорбционна спектрофотометрия, газо-течна хроматография, флуоресцентен анализ), както и радиоимунологични методи.
Соли на жлъчните киселини.
Детергентните ефекти на жлъчката се дължат на жлъчните соли. Стероидната част на молекулата на жлъчната киселина има хидрофобни свойства, характерни за липидите, а окислените странични вериги са хидрофилни. Тази двойна разтворимост, характерна за детергентите и сапуните, се обозначава с термина амфипатичност:
Хидрофобният край на молекулата се смесва лесно с липидите
Хидрофилният край не се смесва лесно с липидите, но улеснява контакта с водната фаза.
Това образува емулсия от липиди във вода, състояща се от малки капчици мазнина.
Образуването на емулсии улеснява тяхното взаимодействие като субстрати с липазите.
Друг вид е образуването на мицели. В този случай хидрофилните части взаимодействат с водната среда, изпъкнали навън. Хидрофилните части остават свързани помежду си във вътрешната сфера на мицела. В мицела обаче няма обширна вътрешна липидна фаза, тъй като нейната дебелина съответства на размера на една молекула. Мицелите са с няколко порядъка по-малки от частиците на емулсията.
Емулгирането е важно, защото увеличава контактната повърхност, когато липазата действа върху мускула, но частиците на емулсията са твърде големи, за да преминат през клетъчната мембрана. Мицелите се абсорбират лесно от епителните клетки на тънките черва.
Структурата на тези мицели е такава, че тяхната хидрофобна сърцевина (мастни киселини, глицериди и др.) е заобиколена отвън от хидрофилна обвивка от мастни киселини и фосфатиди.
Като част от мицелите, висшите мастни киселини и моноглицеридите се прехвърлят от мястото на хидролиза на мазнините към абсорбиращата повърхност на клетъчния епител. По отношение на механизма на абсорбция на мицелите няма консенсус:
1. Вероятно в резултат на мицеларна дифузия и вероятно пиноцитоза мицелите като цяло частица проникват в епителните клетки на вилите. Тук се случва разграждането на мастните мицели, докато жлъчните киселини незабавно навлизат в кръвта и през системата портална венадо черния дроб, откъдето се ресекретира в жлъчката.
2. Само липидният компонент на мастните мицели може да премине в клетките на вилите, докато самите жлъчни киселини, след като са изпълнили своята физиологична функция, остават в чревния лумен. И едва тогава те се абсорбират в кръвта (в илеума), влизат в черния дроб и след това се екскретират с жлъчката.
Триглицеридите и фосфолипидите, ресинтезирани в чревните епителни клетки, както и холестеролът, влизащ в тези клетки, се комбинират с малко количество протеин и образуват относително стабилни сложни частици - хиломикрони. Те са големи по размер и не могат да проникнат в кръвоносните капиляри и дифундират в лимфната система на червата, а от нея в гръдния лимфен канал и по-нататък в кръвообращението, т.е. с тяхна помощ ендогенните триглицериди, холестерол и частично фосфолипиди се транспортират от червата в кръвта.
Вътреклетъчна липидна хидролиза.
Източникът на мастни киселини, които се подлагат на окисление в тъканите на висшите животни, е или извънклетъчна течност, или ендогенни вътреклетъчни липиди. В мускулната тъкан, включително сърдечния мускул, мастните киселини претърпяват директно окисление. Основен източникендогенни мастни киселини - резервни мазнини, съдържащи се в цитоплазмата. Мастните киселини първо се хидролизират от специфични вътреклетъчни липазни ензими до глицерол и свободни мастни киселини, а последните се подлагат на активиране и окисление. Друг източник на мастни киселини са мембранните фосфоглицериди. Постоянно протича метаболитно обновяване на фосфоглицеридите, при което се образуват свободни мастни киселини.
Разграждането на мазнините.
Слюнката не съдържа ензими, разграждащи мазнините. В стомаха също няма забележимо смилане на хранителни мазнини, но има частично разрушаване на липопротеиновите комплекси на мембраните на хранителните клетки, което прави мазнините по-достъпни за последващо излагане на липаза от панкреатичен сок.
Разграждането на мазнините се извършва при хората главно в горните отдели на тънките черва, където има условия за емулгиране на мазнините. Най-мощният емулгиращ ефект върху мазнините се упражнява от жлъчните соли, които навлизат в дванадесетопръстника с жлъчката под формата на натриеви соли. Жлъчните киселини са крайният продукт на метаболизма на холестерола. По своята химическа природа жлъчните киселини са производни на холановата киселина. Жлъчните киселини присъстват в жлъчката в конюгирана форма. Тези връзки понякога се наричат сдвоени, защото. те се състоят от два компонента: жлъчна киселина и глицин и жлъчна киселина и таурин. Жлъчните соли намаляват повърхностното напрежение на мастно-водната повърхност. Жлъчните киселини също играят важна роля като активатор на панкреатичната липаза, под влиянието на която се извършва разграждането на мазнините в червата. Липазата, произведена от панкреаса, разгражда триглицеридите, които са в емулгирано състояние. Чревната липаза също участва в разграждането на мазнините, тя катализира хидролитичното разграждане на моноглицеридите и не действа върху ди- и триглицеридите. По този начин в тънките черва основната част от мазнините се абсорбира след разделяне на мастни киселини и глицерол, които, тъй като са силно разтворими във вода, се абсорбират в червата и след това влизат в кръвния поток на порталната вена и оттам до Черният дроб.
Дълговерижните мастни киселини и моноглицеридите се абсорбират с участието на жлъчката (жлъчни киселини). Мастните киселини и моноглицеридите образуват мицели, които са стабилни във водна среда. Основният ендогенен източник на мастни киселини е резервната мазнина. Триглицеридите от мастното депо играят същата роля в липидния метаболизъм, както чернодробният гликоген играе във въглехидратния метаболизъм. Само свободни мастни киселини могат да се използват като енергийни източници и триглицеридите трябва да бъдат хидролизирани от липази до глицерол и свободни мастни киселини. Те могат да преминат от мастните депа в кръвната плазма и да се използват като енергиен материал.
Друг източник на мастни киселини са мембранните фосфолипиди. Клетките непрекъснато претърпяват метаболитно обновяване на фосфолипидите, по време на което се образуват свободни мастни киселини (фосфолипиди).
Окисляването на мастни киселини в клетките се извършва в митохондриите с участието на мултиензимен комплекс. Процесът на окисление на мастни киселини се състои от следните стъпки:
Етап 1 - активиране на мастни киселини и тяхното проникване от цитоплазмата в митохондриите, с образуване на ацил-КоА. Включва 3 етапа:
а) ензимна естерификация на свободна мастна киселина от цитоплазмения CoA (за сметка на енергията на АТФ) във външната мембрана на митохондриите.
б) прехвърляне на остатък от мастна киселина от CoA към молекула карнитин, с помощта на която този остатък се транспортира през вътрешната митохондриална мембрана.
в) прехвърляне на остатък от мастна киселина от карнитин към интрамитохондриален CoA.
Етап 2 - първият етап на дехидрогениране:. Когато мастните киселини се окисляват, това следва етапа на активиране и се появява вътре в митохондриите. Ацил-КоА претърпява ензимно дехидрогениране, превръщайки се в КоА естер на ненаситена киселина.
Етап 3 - етап на хидратация: добавяне на водна молекула и образуване на b-хидроксил CoA.
Етап 4 - вторият етап на дехидрогениране, образува се b-кетоацил CoA.
Етап 5 - тиолазна реакция: в него се образува ацетил CoA, който претърпява окисление в цикъла на Кребс, и ацил CoA, който след това преминава през b окислителния път. Стъпката на тиолитичното разцепване е тиолиза, силно екзорганична реакция.
Последователното окислително разцепване на ацетил CoA молекули от CoA естери на наситени мастни киселини с дълга верига се нарича b окисление.
Енергиен баланс: В случай на окисление, например палмитинова киселина, G е 9797 kJ.
ЛИТЕРАТУРА
Компоненти на клетките и техните мембрани. Рибозата и дезоксирибозата, захари, съдържащи 5 въглеродни атома, са част от рибонуклеинова (РНК) и дезоксирибонуклеинова (ДНК) киселини. Въглехидратният метаболизъм в човешкото тяло се състои главно от следните процеси: 1. Разграждане в стомашно-чревния тракт до монозахариди, ди- и полизахариди, които идват с храната. Абсорбция в кръвта в червата. ...
Оранжеви тонзили и натрупване на холестеролни естери в други ретикулоендотелни тъкани. Патологията е свързана с ускорен катаболизъм на аро A-I. Храносмилане и усвояване на липиди. Жлъчка. Значение. В зората на формирането на съвременната доктрина за екзокринната функция на черния дроб, когато естествените учени имаха само първите ...




Hanseleit през 1932 г. извежда уравненията за реакциите на синтез на урея, които са представени под формата на цикъл, който в литературата се нарича орнитинов цикъл на образуване на урея на Кребс. Трябва да се отбележи, че в биохимията това е първата циклична метаболитна система, чието описание предшества откриването от Г. Кребс на друг метаболитен процес - цикъла на трикарбоксилната киселина, с почти 5 години. По-нататък...





Клетъчно взаимодействие Външен слой на клетъчните мембрани Стероиди Мембранни компоненти; прекурсори в синтеза на жлъчни киселини и стероидни хормони Клетъчни мембрани; кръвни липопротеини 3. Ролята на липидите в храненето на човека Растителните мазнини и масла са основен компонент на храната, източник на енергия и пластичен материал за човека, доставчик на редица необходими за него ...
БЕЛОРУСКИ ДЪРЖАВЕН УНИВЕРСИТЕТ ПО ИНФОРМАТИКА И РАДИОЕЛЕКТРОНИКА
Катедра ЕТТ
"Основи на липидната биохимия в човешкото тяло"
МИНСК, 2008 г
Липиди е общо наименование за всички известни мазнини и мастоподобни вещества с различна структура, но общи свойства (неразтворимост във вода, екстракция с неорганични разтворители). Липид (гръцка мазнина).
В човешкото тяло 10-20% мазнини от телесното тегло. Липидите са:
Протоплазмен- са част от всички структури на клетките, органите и тъканите и практически остават на едно и също ниво през целия живот. Те съставляват 25% от всички телесни мазнини.
Резервни липиди- съхраняват се в тялото, като броят им варира в зависимост от възрастта, пола, хранителните условия, видовете дейност.
Функции на липидите в организма :
1 – пластична функция: участват в изграждането на клетъчните мембрани на всички органи и тъкани и образуването на много биологично важни съединения (хормони, мастноразтворими витамини).
2 – енергийна функция: липидите осигуряват 25-30% от енергийните нужди на тялото. Разграждането на 1 g мазнини е 9,3 kcal.
3 - мазнините са резервни хранителни вещества, тяхното депо - подкожна тъкан, периренална капсула.
4 – защитенфункцията на липидите: те участват в терморегулацията, предпазват кожата от изсушаване, органите от сътресения.
5- изпълнява функцията защитни черупки, предпазвайки от инфекция или прекомерна загуба или натрупване на вода.
6 - осигуряват усвояването на мастноразтворимите витамини
Класификация на липидите:
1 - простоили неутрални мазнини (естери на мастни киселини и алкохоли).Неутралните мазнини са в тялото или под формата на протоплазмена мазнина, която е структурен компонент на клетките, или под формата на резервна, резервна мазнина.
2 - сложни мазнини, са естери на тривалентен алкохол глицерол, високомолекулни мастни киселини и други компоненти. Сред сложните мазнини са: фосфолипиди, гликолипиди, сфигномиелини. Сфинголипидите се намират в мембраните на животински и растителни клетки.
3 - липидни производни. Те включват всички съединения, които не могат да бъдат ясно класифицирани като прости или сложни липиди, като стероиди, каротеноиди и витамини от липиден характер.
4- восък- например ланолин, смес от холестеролови естери.
восък -Това са естери, образувани от наситени и ненаситени мастни киселини и алкохоли.
Намира се в неутрални мазнини :
Мастна киселина.
Мастните киселини получават името си от начина, по който са изолирани от мазнините. Това са карбоксилни киселини с дълга алифатна верига.
Естествените мастни киселини са много разнообразни. Повечето мастни киселини са монокарбоксилни киселини, съдържащи линейни въглехидратни вериги с четен брой атоми. Съдържанието на ненаситени мастни киселини е по-високо от това на наситени. Ненаситените мастни киселини имат по-ниска точка на топене.
свойства на мастните киселини.
Наситените и ненаситените мастни киселини се различават значително по своята структурна конфигурация. В наситените мастни киселини въглеводородната опашка може по принцип да приеме различни конформации поради пълната свобода на въртене около крайната единична връзка.
При ненаситените киселини се наблюдава различна картина: невъзможността за въртене около двойната връзка осигурява твърд завой във въглеводородната верига.
Естествените мастни киселини, наситени и ненаситени, не абсорбират светлина нито във видимата, нито в UV област. Спектрофотометрично се определя само след изомеризация (230-260 nm). Ненаситените се определят чрез количествено титруване. Анализът на сложни смеси от мастни киселини се извършва чрез газово-течна хроматография.
Наситен-палмитинова, стеаринова, липоцеринова киселини
Ненаситени: арахидонова, олеинова, линолова, линоленова.
Растителните мазнини се състоят главно от ненаситени мастни киселини.
Липидите са съществена част от балансираната човешка диета. Съотношението на протеини, липиди и въглехидрати трябва да бъде 1:1:4.
Стойността на мазнините е много разнообразна. Високото им съдържание на калории им придава специална стойност. Мазнините са разтворители на витамини А, D, Е и др. С мазнините в организма се въвеждат някои ненаситени киселини, които се класифицират като есенциални мастни киселини (линолова, линоленова, арахидонова), които не се синтезират при хора и животни. С мазнините в тялото влиза комплекс от биологично активни вещества: фосфолипиди, стероли.
ТриацилглицеролиОсновната им функция е да съхраняват липиди. Те се намират в цитозола под формата на фини емулгирани маслени капчици.
Сложни мазнини :
Фосфолипиди- основните компоненти на клетъчните мембрани и субклетъчните органели, изграждат по-голямата част от тъканите на мозъка, нервите, черния дроб, сърцето, участват в биосинтезата на протеини, активирането на протромбина, транспортирането на липиди и мастноразтворими витамини в кръвта и лимфата. Състои се от глицерол и две молекули мастни киселини, едната от които е наситена. а другата е ненаситена + азотна основа.
Липопротеини .
Полярните липиди се свързват с определени специфични протеини, за да образуват липопротеини, от които транспортните липопротеини са най-известни и присъстват в кръвната плазма на бозайниците.
В такива сложни липиди взаимодействията между липидите и протеиновите компоненти се осъществяват без участието на ковалентни връзки.
Липопротеините обикновено съдържат както полярни, така и неутрални липиди, както и холестерол и неговите естери. Те служат като формата, в която липидите се транспортират от тънките черва до черния дроб и от черния дроб до мастната тъкан и други тъкани.
В кръвната плазма са открити няколко класа липопротеини, като тяхната класификация се основава на разликите в тяхната плътност. Липопротеини с различни съотношения на липиди и протеини могат да бъдат разделени в ултрацентрофуга.
Най-леките липопротеини са хиломикроните: големи структури, съдържащи около 80% триацилглицероли, 7% фосфоглицериди, 8% холестерол и неговите естери и 2% протеин.
Плазмените бета-липопротеини съдържат 80-90% липиди, а алфа-липопротеините - 40-70%.
Точната структура на липопротеините все още не е известна, но има причини да се смята, че протеиновата верига е разположена на външната повърхност, където образува тънка хидрофилна обвивка около мицеларната липидна структура. Мазнините или триглицеридите съхраняват по-голямата част от енергията, освободена в резултат на химични реакции.
Наред с неполярните липиди има полярни липиди. Те са основните компоненти на клетъчните мембрани. Множество ензими и транспортни системи са локализирани в мембраните. Много свойства на клетъчните мембрани се дължат на наличието в тях на полярни липиди.
Мембранни липиди:
Мембранните липиди, заедно с въглеводородните вериги, съдържат една или повече силно полярни „глави“. Фосфолипидите присъстват в малки количества в мембраните. Техният основен компонент - фосфоглицеридите - съдържат 2 мастни киселинни остатъка, които естерифицират първата и втората хидроксилна група на глицерола. Третата хидроксилна група образува естерна връзка с фосфорната киселина. Хидролизира при нагряване с киселини и основи, както и ензимно - под действието на фосфолипази.
Сфинголипидите са вторият клас мембранни липиди, те имат полярна глава и две неполярни опашки, но не съдържат глицерол.
Те са разделени на 3 подкласа: сфингомиелини, цереброзиди и ганглеозиди.
Сфингомиелините се намират в миелиновите обвивки на някои видове нервни клетки. Церброзиди – в мембраните на мозъчните клетки. Ганглеозидите са важни компоненти на специфични рецепторни места, разположени на повърхността на клетъчните мембрани. Те се намират в онези специфични области на нервните окончания, където се осъществява свързването на невротрансмитерни молекули в процеса на химическо предаване на импулс от една нервна клетка към друга.
Изследвани са външните или плазмените мембрани на много клетки, както и мембраните на редица вътреклетъчни органели, като митохондрии и хлоропласти. Всички мембрани съдържат полярни липиди.
Липидната част на мембраната е смес от полярни липиди. Естествените мембрани се характеризират с малка дебелина (6-9nm) и еластичност. Водата лесно преминава през мембраните, но те са практически непропускливи за замърсени йони като натрий, хлор или водород и за полярни, но незамърсени захарни молекули. Полярните молекули проникват с помощта на специфични носители на транспортната система.
Фосфоглицеридите, сфинголипидите, гликолипидите и восъците често се наричат осапуняеми липиди, тъй като при нагряването им (в резултат на елиминирането на мастни киселини) се образуват сапуни. Клетките също така съдържат по-малко неосапуняеми липиди, те не се хидролизират с освобождаване на мастни киселини.
Има 2 вида неосапуняеми липиди:
Стероиди и терпени
Стероиди - жлъчни киселини, полови хормони, надбъбречни хормони.
Стероидите са широко разпространени в природата. Тези съединения включват множество вещества от хормонален характер, както и холестерол, жлъчни киселини и др.
Стероли – Холестерол Холестеролът играе ролята на междинен продукт в синтеза на много други съединения. Холестеролът е богат на плазмените мембрани на много животински клетки; той се намира в много по-малки количества в мембраните на митохондриите и в ендоплазмения ретикулум.
Растенията съдържат фитостероли.
Терпени - намират се в растенията, много от тях придават на растенията характерния аромат и служат като основни компоненти на "ароматните масла".
мастноразтворими витамини.
Витамините са жизненоважни вещества, присъстващи в тялото в следи и необходими за нормалните клетъчни функции. Мастноразтворимите витамини включват витамини А, Е, К, D, молекулярната основа на тяхното действие не е точно известна.
Витамините се класифицират като липиди, защото са неразтворими във вода и могат да се екстрахират с органични разтворители. Мастноразтворимите витамини имат изопреноидна структура (A, E, K), витамин D е стероидно производно, въпреки че стероидите също идват от изопреноидни прекурсори. Витамин А се намира само в животинските тъкани. Съществува в 2 химични форми А-1 и А-2 (витамири) - ретинол 1 и ретинол 2 и представлява алкохол, съдържащ ацикличен пръстен, към който е прикрепена странична верига, състояща се от 2 изопренови единици.
Растенията съдържат каротеноидни пигменти. Алфа-, бета- и гама-каротините, при окислителното им разграждане в животинските тъкани се образува витамин А. Липсата на витамин А води до нарушен растеж и развитие на „нощна слепота“, нарушава се нормалната функция на пръчките на ретината.
Витамин ЕТой е представен от цяла група витамини, присъстващи в растителните масла и наречени токофероли. Тези съединения имат ароматна пръстенна система, съдържаща хидроксил, и изопреноидна странична верига. Недостигът води до атрофия и мускулна слабост, стерилитет. Смята се, че тези вещества предотвратяват разрушителното действие на молекулярния кислород, понякога се наричат антиоксиданти.
Витамин К– K 1 и K 2, нафтохинони с дълги странични изопреноидни вериги с различна дължина. Недостигът се проявява в нарушение на процеса на съсирване на кръвта поради загубата на способността на организма да синтезира протромбин.
Витамин D- производно на стероиди. Най-важният е витамин D 2 - калциферол и D 3. Липсата на витамин D води до нарушения на метаболизма на калция и фосфора, което се проявява в промени в структурата на костите и зъбите. Витамин D насърчава абсорбцията на Ca йони в тънките черва чрез стимулиране на синтеза на протеин, участващ в транспорта на тези йони.
Простагландини.
Тези вещества се намират в почти всички органи и тъкани на хора и животни, тяхната висока активност, широк спектър на действие е сравнима с ефекта на хормоните.
Простагландините са циклични полиненаситени мастни киселини с относително молекулно тегло 300-400. Те съдържат само въглерод, кислород и водород.
Биосинтезата на простагландини се осъществява в микрозомите на клетката. Ненаситените мастни киселини са предшественици на простагландините. Те се синтезират при физиологична нужда. Тяхната роля е най-активните хранителни вещества. Те намаляват стомашната секреция, засягат гладката мускулатура, сърдечно-съдовата система.
Те се определят чрез абсорбционна спектрофотометрия, газо-течна хроматография, флуоресцентен анализ), както и радиоимунологични методи.
Соли на жлъчните киселини.
Детергентните ефекти на жлъчката се дължат на жлъчните соли. Стероидната част на молекулата на жлъчната киселина има хидрофобни свойства, характерни за липидите, а окислените странични вериги са хидрофилни. Тази двойна разтворимост, характерна за детергентите и сапуните, се обозначава с термина амфипатичност:
Хидрофобният край на молекулата се смесва лесно с липидите
Хидрофилният край не се смесва лесно с липидите, но улеснява контакта с водната фаза.
Това образува емулсия от липиди във вода, състояща се от малки капчици мазнина.
Образуването на емулсии улеснява тяхното взаимодействие като субстрати с липазите.
Друг вид е образуването на мицели. В този случай хидрофилните части взаимодействат с водната среда, изпъкнали навън. Хидрофилните части остават свързани помежду си във вътрешната сфера на мицела. В мицела обаче няма обширна вътрешна липидна фаза, тъй като нейната дебелина съответства на размера на една молекула. Мицелите са с няколко порядъка по-малки от частиците на емулсията.
Емулгирането е важно, защото увеличава контактната повърхност, когато липазата действа върху мускула, но частиците на емулсията са твърде големи, за да преминат през клетъчната мембрана. Мицелите се абсорбират лесно от епителните клетки на тънките черва.
Структурата на тези мицели е такава, че тяхната хидрофобна сърцевина (мастни киселини, глицериди и др.) е заобиколена отвън от хидрофилна обвивка от мастни киселини и фосфатиди.
Като част от мицелите, висшите мастни киселини и моноглицеридите се прехвърлят от мястото на хидролиза на мазнините към абсорбиращата повърхност на клетъчния епител. По отношение на механизма на абсорбция на мицелите няма консенсус:
1. Вероятно в резултат на мицеларна дифузия и вероятно пиноцитоза мицелите като цяло частица проникват в епителните клетки на вилите. Тук се разграждат мастните мицели, докато жлъчните киселини веднага навлизат в кръвния поток и през системата на порталната вена в черния дроб, откъдето отново се секретират като част от жлъчката.
2. Само липидният компонент на мастните мицели може да премине в клетките на вилите, докато самите жлъчни киселини, след като са изпълнили своята физиологична функция, остават в чревния лумен. И едва тогава те се абсорбират в кръвта (в илеума), влизат в черния дроб и след това се екскретират с жлъчката.
Триглицеридите и фосфолипидите, ресинтезирани в чревните епителни клетки, както и холестеролът, влизащ в тези клетки, се комбинират с малко количество протеин и образуват относително стабилни сложни частици - хиломикрони. Те са големи по размер и не могат да проникнат в кръвоносните капиляри и дифундират в лимфната система на червата, а от нея в гръдния лимфен канал и по-нататък в кръвообращението, т.е. с тяхна помощ ендогенните триглицериди, холестерол и частично фосфолипиди се транспортират от червата в кръвта.
Вътреклетъчна липидна хидролиза .
Източникът на мастни киселини, които се подлагат на окисление в тъканите на висшите животни, е или извънклетъчна течност, или ендогенни вътреклетъчни липиди. В мускулната тъкан, включително сърдечния мускул, мастните киселини претърпяват директно окисление. Основният източник на ендогенни мастни киселини са резервните мазнини, съдържащи се в цитоплазмата. Мастните киселини първо се хидролизират от специфични вътреклетъчни липазни ензими до глицерол и свободни мастни киселини, а последните се подлагат на активиране и окисление. Друг източник на мастни киселини са мембранните фосфоглицериди. Постоянно протича метаболитно обновяване на фосфоглицеридите, при което се образуват свободни мастни киселини.
Разграждането на мазнините.
Слюнката не съдържа ензими, разграждащи мазнините. В стомаха също няма забележимо смилане на хранителни мазнини, но има частично разрушаване на липопротеиновите комплекси на мембраните на хранителните клетки, което прави мазнините по-достъпни за последващо излагане на липаза от панкреатичен сок.
Разграждането на мазнините се извършва при хората главно в горните отдели на тънките черва, където има условия за емулгиране на мазнините. Най-мощният емулгиращ ефект върху мазнините се упражнява от жлъчните соли, които навлизат в дванадесетопръстника с жлъчката под формата на натриеви соли. Жлъчните киселини са крайният продукт на метаболизма на холестерола. По своята химическа природа жлъчните киселини са производни на холановата киселина. Жлъчните киселини присъстват в жлъчката в конюгирана форма. Тези връзки понякога се наричат сдвоени, защото. те се състоят от два компонента: жлъчна киселина и глицин и жлъчна киселина и таурин. Жлъчните соли намаляват повърхностното напрежение на мастно-водната повърхност. Жлъчните киселини също играят важна роля като активатор на панкреатичната липаза, под влиянието на която се извършва разграждането на мазнините в червата. Липазата, произведена от панкреаса, разгражда триглицеридите, които са в емулгирано състояние. Чревната липаза също участва в разграждането на мазнините, тя катализира хидролитичното разграждане на моноглицеридите и не действа върху ди- и триглицеридите. По този начин в тънките черва основната част от мазнините се абсорбира след разделяне на мастни киселини и глицерол, които, тъй като са силно разтворими във вода, се абсорбират в червата и след това влизат в кръвния поток на порталната вена и оттам до Черният дроб.
Дълговерижните мастни киселини и моноглицеридите се абсорбират с участието на жлъчката (жлъчни киселини). Мастните киселини и моноглицеридите образуват мицели, които са стабилни във водна среда. Основният ендогенен източник на мастни киселини е резервната мазнина. Триглицеридите от мастното депо играят същата роля в липидния метаболизъм, както чернодробният гликоген играе във въглехидратния метаболизъм. Само свободни мастни киселини могат да се използват като енергийни източници и триглицеридите трябва да бъдат хидролизирани от липази до глицерол и свободни мастни киселини. Те могат да преминат от мастните депа в кръвната плазма и да се използват като енергиен материал.
Друг източник на мастни киселини са мембранните фосфолипиди. Клетките непрекъснато претърпяват метаболитно обновяване на фосфолипидите, по време на което се образуват свободни мастни киселини (фосфолипиди).
Окисляването на мастни киселини в клетките се извършва в митохондриите с участието на мултиензимен комплекс . Процесът на окисление на мастни киселини се състои от следните стъпки:
Етап 1 - активиране на мастни киселини и тяхното проникване от цитоплазмата в митохондриите, с образуване на ацил-КоА. Включва 3 етапа:
а) ензимна естерификация на свободна мастна киселина от цитоплазмения CoA (за сметка на енергията на АТФ) във външната мембрана на митохондриите.
б) прехвърляне на остатък от мастна киселина от CoA към молекула карнитин, с помощта на която този остатък се транспортира през вътрешната митохондриална мембрана.
в) прехвърляне на остатък от мастна киселина от карнитин към интрамитохондриален CoA.
Етап 2 - първият етап на дехидрогениране:. Когато мастните киселини се окисляват, това следва етапа на активиране и се появява вътре в митохондриите. Ацил-КоА претърпява ензимно дехидрогениране, превръщайки се в КоА естер на ненаситена киселина.
Етап 3 - етап на хидратация: добавяне на водна молекула и образуване на b-хидроксил CoA.
Етап 4 - вторият етап на дехидрогениране, образува се b-кетоацил CoA.
Етап 5 - тиолазна реакция: в него се образува ацетил CoA, който претърпява окисление в цикъла на Кребс, и ацил CoA, който след това преминава през b окислителния път. Стъпката на тиолитичното разцепване е тиолиза, силно екзорганична реакция.
Последователното окислително разцепване на ацетил CoA молекули от CoA естери на дълговерижни наситени мастни киселини се нарича b окисление.
Енергиен баланс: В случай на окисление, например палмитинова киселина, G е 9797 kJ.
Най-често срещаните киселини в зеленчуците, горските плодове и плодовете са нелетливи лимонена, ябълчена, винена и други киселини. Съдържанието на киселини в дивите плодове варира от 0,6 до 6%. Най-високата киселинност е характерна за боровинките, морския зърнастец, калината.
Киселините участват във формирането на вкуса на дивите плодове, намаляват pH на околната среда, влияят благоприятно на процеса на храносмилане, допринасят за образуването на определен състав на микрофлората и инхибират процесите на гниене в стомашно-чревния тракт. Фенолните киселини имат бактерициден ефект.
В допълнение към нелетливите киселини, плодовете и плодовете съдържат малко количество летливи киселини: мравчена, валерианова и капронова. Всички те се съдържат в състава на калината. Сорбиновата и парасорбиновата киселина са характерни за плодовете на планинската пепел. Мравчена киселина се съдържа в малините. Фенолните киселини и техните естери се намират в малки количества в различни плодове: например салициловата киселина се намира в малини, къпини, ягоди, боровинки, планинска пепел. Летливите киселини формират аромата на пресни плодове.
Липиди.Съставът на плодовете и плодовете съдържа някои липиди
(0,1-0,3%), те са концентрирани главно в семената. Изключение прави морският зърнастец, който съдържа липиди както в пулпата, така и в семената. Концентрацията на липиди в семената варира от 4 до 31%.
Липидите играят важна роля, тъй като са част от клетъчните мембрани. В плодовете и горските плодове липидите са представени от мазнини, восъци, кутин, стероиди и други съединения. Мазнините от плодове и плодове се характеризират с благоприятен мастнокиселинен състав с преобладаване на ненаситени мастни киселини.
Восъците, кутинът, урсоловата киселина образуват восъчно покритие, характерно например за боровинките и червените боровинки.
витамини. Хранителната стойностзеленчуци, плодове и плодове до голяма степен се определя от наличието на витамини и витаминоподобни вещества в тях. Плодовете и горските плодове съдържат вода и мастноразтворими витамини. Водоразтворимите включват аскорбинова киселина, витамини от група В, биофлавоноиди.
Витамин С, по-точно неговата редуцирана форма - L-аскорбинова киселина, се среща често в дивите плодове и горски плодове. Много от тях при редовна употреба могат лесно да запълнят дневната нужда от аскорбинова киселина (50-100 mg на ден).
Аскорбиновата киселина е активен участник в много редокс процеси, протичащи в човешкото тяло. Подобрява устойчивостта към инфекциозни заболявания, особено настинки, неблагоприятни външни влияния (прегряване, охлаждане), повишава ефективността. Аскорбиновата киселина има ефект върху хематопоезата, метаболизма на въглехидратите и холестерола. Биологичната роля на витамин С се проявява най-пълно в присъствието на Р-активни съединения, което е характерно за плодовете и горските плодове. За човек дневната норма на аскорбинова киселина е 70-100 mg, P-активни вещества - 30-50 mg.
Плодовете от семейството на червените боровинки, както и къпините, боровинките, черешите съдържат 10-20 mg витамин С / 100 грама.Средното му количество (от 30 до 75 mg / 100 g) е характерно за ягоди, калина, планинска пепел, боровинки.
Касисът по съдържание на аскорбинова киселина надминава много видове плодове, натрупвайки го до 500 mg / 100 грама.
Групата на Р-активните съединения включва антоцианини, левкоантоцианини, катехини, флавоноли, които се различават по химичен състав. Р-активните съединения имат антихипертензивен и антисклеротичен ефект. Общото съдържание на P-активни вещества е 0,-0,6% в плодовете и плодовете. В интензивно оцветените и аронийните култури, като боровинки, касис, боровинки, съдържанието на Р-активни вещества достига 1-1,5%.
Витамините от група В са включени в простатната група ензими, които извършват енергийния метаболизъм, биосинтезата на пуринови основи; те регулират метаболизма на въглехидратите, протеините и мазнините. От дивите плодове източникът на рибофлавин са шипките, боровинките, боровинките, а източникът на тиамин е планинската пепел.
Голямото и умерено съдържание на рибофлавин е характерно за зеления грах, сладките пиперки и карфиола.
При липса на тиамин и рибофлавин се развива авитаминоза, проявяваща се в повишена умора.
Представители на семейството на витамин B6 са открити в много горски плодове.- пиридоксин, пиридоксал, пиридоксамин, но в малки количества (от 0,1 до 3,9 mg / 100 грама). Физиологичната нужда на възрастен от този витамин е 1,8-2 mg на ден. Недостигът му води до нервни разстройства, сънливост, възпаление на кожата, очите, докато нуждата от ниацин се увеличава, използването на аминокиселини и протеини се нарушава.
Ниацин (PP) се изисква от човек в значителни количества - 14-28 mg на ден. При липсата му в диетата се появяват раздразнителност, депресивно настроение, главоболие и други болезнени симптоми. Ниацинът се съдържа във всички горски плодове. Боровинките, шипките, морският зърнастец и малините съдържат повече ниацин от останалите.
Източникът на витамини от група В и РР също са картофи, карфиол, гъби - манатарки (28,15 мкг / 100 г) и лисички (25,38 мкг / 100 г).
Някои плодове са най-ценни като източник на фолиева киселина.Този витамин е антианемичен фактор, стимулира образуването на червени и бели кръвни клетки. От плодовете, които могат да задоволят нуждата на човек от този витамин, е необходимо да се отбележи планинска пепел, дива роза, ягоди и малини.
От зеленчуците фолиевата киселина се намира в зеления грах, зелето (20-40 mcg на 100 g продукт), а от гъбите - в белите гъби, манатарките и шампиньоните (40 mcg; съответно 30 mcg и 30 mcg).
Мастноразтворимите витамини включват A, D, E, K. Горските плодове с наситено жълта плът са богати на каротеноиди, включително В-каротин, който най-ефективно се превръща във витамин А в човешкото тяло.Основните акумулатори на В-каротин са морски зърнастец, глог, шипка. Каротеноидите предотвратяват тъканната хепаксия, участват в образуването на надбъбречен хормон, зрителен пигмент.
Източникът на тези витамини са зеленчуци като карфиол, моркови и домати.
Витамин Е стабилизира и предпазва ненаситените липиди от прекомерно окисляване и предотвратява безплодието при хората. Най-високата биологична активност е характерна за а-токоферола. Токоферолът се синтезира само от растения. Облепихата е изключително богата на тях – до 18 mg/100 g в плодовете и десет пъти повече в маслото. Шипка, глог, планинска пепел са богати на токофероли.
Витамин К подпомага съсирването на кръвта. В плодовете се съдържа под формата на К1. Повече от другите плодове витамин К1 се натрупва от планинска пепел, касис, дива роза, боровинка.
ЛИПИДИ
Липидите (от гръцки - мастни) са естествени неполярни съединения, които са напълно или почти напълно неразтворими във вода, но разтворими в неполярни разтворители, например хлороформ, въглероден дисулфид, етер и горещ етанол. Основната отличителна черта на тази група от доста хетерогенни вещества е тяхната неполярност, която в някои случаи се обяснява с наличието в тези съединения на една или повече мастни киселини, съдържащи дълги алифатни въглеводородни вериги. Съдържанието на липиди в организма е 10 - 20% от телесното тегло.
Липидите изпълняват следните биологични функции :
1. Те са компоненти на мембраните, които до голяма степен определят тяхната пропускливост.
2. Те служат като една от основните форми на съхранение на въглерод и енергия (1 g мазнини по време на окисление дава 9,3 kcal).
3. Могат да бъдат прекурсори на други важни съединения.
4. Изпълняват ролята на защитни бариери, които предпазват от топлинни, електрически и физически въздействия.
5. Те са част от защитни обвивки, които предпазват от инфекция и прекомерна загуба или натрупване на вода. Мазнините са източници на ендогенна вода.
6. В някои случаи това са витамини и хормони.
7. Участват в намаляването на нервните импулси, тъй като са част от миелиновите обвивки.
Съществува класификация, базирана на съдържанието на един или друг алкохол в състава на мазнината. Съответно се изолират глицерин-съдържащи, сфингозин-съдържащи липиди и холестерол и неговите производни.
Съществува класификация на липидите според тяхната структура:
1. Прости липиди. 2. Сложни липиди. 3. Липидни производни.
Простите мазнини включват само естери на мастни киселини и алкохоли. Съставът на комплексните липиди, освен мастни киселини и алкохоли, включва и други компоненти. Сред сложните липиди се разграничават фосфоацилглицероли, сфингомиелини, цереброзиди и ганглиозиди. Липидните производни включват всички съединения, които не могат да бъдат ясно класифицирани като прости или сложни липиди, като стероиди, каротеноиди и витамини от липиден характер. Много от тях се образуват от въглеводородни вериги, които са част от мастни киселини.Съставът на простите и сложните липиди задължително включва мастни киселини.Всъщност това са карбоксилни киселини с дълга алифатна верига. Естествените мастни киселини са много разнообразни.
За удобство се разграничават следните групи: 1. Повечето мастни киселини са монокарбоксилни киселини, съдържащи линейни въглеводородни вериги с четен брой атоми (обикновено C 12 - C 20). По-рядко срещани са ФК с по-къси вериги или с нечетен брой въглеродни атоми. 2. Често има киселини, съдържащи етиленови (ненаситени, двойни) връзки (обикновено C 18 - C 20 киселини). Ако има две или повече двойни връзки, те почти винаги са разделени от една метиленова група: - CH = CH - CH 2 - CH = CH -.
3. В ненаситените киселини с двойни връзки почти винаги има цис конфигурация.
Голям брой неполярни C - C и C - H връзки във въглеродната верига придават по същество неполярен характер на цялата молекула като цяло, въпреки че тя също има полярна група - COOH. Съвсем ясно е, че съединение, съдържащо един или повече FA остатъци, ще бъде неполярно, което е причината за неразтворимостта на липидите във вода и също така причинява сглобяването на липидите в биомембрани.
Основните естествени мастни киселини, които изграждат тъканите:
Наситени LCD дисплеи:
1. Лаурин (C 12). 2. Миристик (C 14). 3. Палмитинова (C 16).
4. Стеаринова киселина (C 18). 5. Арахиноична (C 20).
II. Наситени LCD дисплеи:
1. Палмитолеинов (С 16:1) ∆9. 2.Олеинов (С 18:1) ∆9. 3. Линолова (С 18:2) ∆9, 12
4. Линолова (С 18:3) ∆9, 12, 15. 5. Арахидонова (С 20:4) ∆5, 8, 11, 14
Където (С x:y) ∆ Z- x е броят на въглеродните атоми. y е броят на двойните връзки.
Z е позицията на двойните връзки (броят идва от - COOH).
ФК могат да бъдат окислени ензимно и неензимно.
Неензимно окисление на мастни киселини. (LPO) - при двойни връзки те се окисляват до хидроперокси киселини под действието на силни окислители, например H 2 O 2; Около 2; ТОЙ. Получените пероксидни съединения са токсични форми на кислородни съединения. Сложните липиди, присъстващи в биологичните мембрани, много лесно образуват пероксиди, тъй като съдържат много ненаситени мастни киселини. Това от своя страна може да доведе до окисление на мембранните протеини, което може да има дълбок ефект върху структурата и функцията на мембраната. Такъв разрушителен ефект се причинява на първия етап от превръщането на липидните пероксиди в пероксидни радикали - силно реактивни агенти.Тази реакция може да протече с участието на метален йон, например желязо. Гранясването и развалянето на хранителните продукти отчасти се дължат на тези реакции Живите клетки са в състояние да се защитят срещу действието на тези оксиданти. Неутрализирането става с участието на глутатион, витамини Е и С, които действат като антиоксиданти Ензимно окисление на мастни киселини - това е окисление на мастни киселини с дълга верига, по-специално на арахидонова киселина, предшественик на синтеза на простагландин. Най-общо се наричат съединения, образувани от С 20 - полиненаситени киселини ейкозаноиди. Арахидоновата киселина се образува от мембранни фосфолипиди, които под действието на фосфолипаза А2 се разлагат на лизофосфолипиди и арахидонова киселина. Линоловата киселина може също да бъде прекурсор на арахидоновата киселина. Простаглидините (PG) получиха името си, защото за първи път бяха изолирани от семенната плазма, произведена в простатната жлеза. Сега е известно, че те присъстват в повечето тъкани на бозайници, но в изключително малки количества (≤10 -9 g).
Простагландините регулират следните процеси в клетките :
1. Намаляване гладка мускулатура.
2. Кръвообращение (кръвно налягане).
3. Предаване на нервни импулси.
4. Участват във възпалителния процес.
5. Водно-електролитен баланс.
6. Съсирване на кръвта.
Механизмът на действие на простагландините е свързан с наличието в таргетните клетки, контролирани от простагландини, на специфични мембранни протеини - рецептори, които се свързват с определени простагландини. Крайният ефект се медиира от действието на циклични нуклеотиди - простагландините повишават или намаляват (в зависимост от естеството на тъканта) съдържанието на вътреклетъчния cAMP или cGMP. Има 6 основни типа PG. Общ структурна формулапростагландини показва, че това е С20 монокарбоксилна киселина с циклопентанов пръстен вътре във веригата. Различни простагландини имат една или две двойни връзки в специфични позиции и също се свързват в специфични позиции с кислородни атоми.
Биосинтезата на простагландините започва с превръщането на арахидоновата киселина в два ендопероксидни междинни продукта - PGG 2 (образува се първи) PGH 2 . И двете реакции се катализират от един и същ ензим - простагландин ендопероксид синтаза, който се състои от два компонента - циклооксигеназа (присъединяване на O 2 и образуване на циклопентанов пръстен) и пероксидаза (отговорна за образуването на хидропероксидни групи). В някои клетки PGG 2 и PGH 2 се превръщат в други продукти, тромбоксани и простоциклин. Образуването на тромбоксан се случва в тромбоцитите и насърчава съсирването на кръвта, причинявайки агрегация на тромбоцитите и стесняване на артериите. Напротив, образуването на простоциклин се случва в клетките, покриващи артериите и вените, и предотвратява съсирването на кръвта чрез инхибиране на същите процеси. Дисбалансът на тези съединения в полза на агрегацията на тромбоцитите може да бъде основната причина за образуването на адсорбиращи плаки. Биохимията на простагландините обяснява терапевтичния ефект на аспирина, който служи като ефективно средство за облекчаване на лека болка, възпаление, като антипиретик, т.е. състояния, чието възникване до известна степен зависи от повишения синтез на простагландини. Аспиринът намалява скоростта на техния синтез, действайки като инхибитор на циклооксигеназния компонент на ендопероксидния синтез, като същевременно настъпва ковалентна модификация на важната странична аминогрупа на ензима. Друго противовъзпалително средство, индометозин, също инхибира синтеза, но не чрез ковалентна модификация. Лекарствата от стероидна природа действат по различен начин, например чрез инхибиране на образуването на арахидонова киселина от мембранните липиди. Наскоро беше открит нов клас съединения, наречени левкотриени. Те се синтезират от левкоцитите и съдържат три конюгирани двойни връзки. Това са мощни активатори на съкращението на гладката мускулатура. Малките бронхиоли са особено чувствителни към тях, което вероятно се дължи на затрудненото дишане на пациентите бронхиална астма. Ревматоидният артрит също се дължи на възпалителния ефект на левкотриените. Синтезът на левкотриените също започва с арахидонова киселина, но в този случай тя се влияе от ензима липоксигеназа, който превръща арахидоновата киселина в хидропероксид. Нито аспиринът, нито индометоцинът влияят на този ензим. Най-мощните левкотриени са съединения с тиоетерна връзка: - C - S - C - образувани в резултат на кондензация с глутатион.
Помислете за различните видове липиди:
I. Прости липиди .
Простите липиди се разделят на две групи:
1. Неутрални ацилглицероли.
1. Неутрални ацилглицероли.
В зависимост от броя на FA остатъците, свързани с тривалентния алкохол глицерол, се разграничават моно-, ди-, триацил-глицероли. Триацилните производни са най-често срещаните в природата. Във всички случаи простият ацилглицерол не съдържа функционални йонни групи и принадлежи към неутралните липиди. Ацилните странични вериги обикновено са различни.
Обща формула:
CH 2 O - C - R
CHO - C - R /
CH 2 O - C - R //
TAG изпълняват три основни функции в животинските тъкани:
1. В мастната тъкан те се отлагат в резерв (като форма на съхранение на енергия и въглерод). Това са основните липиди на резервните мазнини, има до 99% от тях.
2. под формата на липопротеинови частици (например хиломикрони) се транспортират по лимфна системаи кръвния поток, разпределяйки се до всички тъкани (в този случай тези TAG, които са били синтезирани от усвоени мастни киселини, се транспортират).
3. TAG действат като физическа защита и регулатор на температурата за различни органи на тялото. Най-често (50%) съдържат олеинова киселина, 23% - палмитинова, 10% - линолова, 6% - стеаринова, 5% - палмитоолеинова и 6% - останалите мастни киселини.
TAG се характеризират с мастни киселини с четен брой въглеродни атоми, но има и мастни киселини с нечетен брой въглеродни атоми - те са 2-5%, а кожните мазнини имат 15-20%. Наситеността определя точката на топене на мазнините. Подкожната мазнина е по-наситена от чернодробната мазнина. Съставът на мастните киселини на липидите може да варира от T 0 среда.
2.Восъци.
Това са естери, но както алкохолът (до С18), така и киселината, включени в състава им, имат дълги въглеводородни вериги. Всички восъци са напълно неразтворими във вода. Естествените восъци обикновено са крайните продукти на тези метаболитни пътища, чиято основна роля е да образуват защитни покрития. Птичите пера и животинските кожи имат восъчно покритие, което ги прави водоотблъскващи. Восъчното покритие на листата и плодовете на растенията намалява загубата на влага и намалява възможността от инфекция.
II Комплексни липиди.
Има три основни класа сложни липиди:
1. Фосфоацилглицероли. 2. Сфингомиелини. 3. Гликолипиди.
Първите два класа се наричат фосфатиди или фосфолипиди поради наличието на фосфатни групи. Всички тези съединения се намират само в мембраните, като техни основни компоненти.
1. Фосфоацилглицероли (FAG)
Това са най-често срещаните сложни липиди. Броят на различните ФК и тяхната относителна концентрация варира значително в зависимост от вида на клетките и тяхното физиологично състояние. Въпреки това разнообразие, те все още правят разлика между главни и второстепенни компоненти. Основен:фосфатилхолин (лецитин), фосфатидилетаноламин (цефалин), фосфатидилглицерин. Незначителен:фосфатидилсерин, дифосфатидилглицерол (кардиолипид), фосфатидилинозитол и фосфатидна киселина. Всички други FAGs се образуват от фосфатидна киселина.
CH 2 O - C - R
CHO - C - R 1
CH 2 O - R - R 1
Фосфатна киселина.
Различни FAG се различават един от друг по допълнителни групи, прикрепени чрез фосфоестерна връзка към фосфатидната киселина. Съставът на FA на различни FAG се различава дори в рамките на един и същи организъм. Някои лецитини, като дипалмитоиллецитин, са основният елемент на сърфактанта (повърхностно активното вещество) на белите дробове - липсата му при недоносени бебета причинява дихателна недостатъчност.
Структура на някои ФА:
CH 2 O - C - R CH 2 O - C - R
CHO - C - R1 CHO - C - R 1
CH 2 O - R - OH CH 2 O - R - OH
O - CH 2 - CH 2 - N + H 3 O - CH 2 - CH 2 - N + - (CH 3) 3
Фосфатидилетаноламин Фосфатидилхолин
(цефалин) (лецитин)
CH 2 O - C - R CH 2 O - C - R
CHO - C - R 1 CHO - C - R 1
CH 2 O - R - OH N + H 3 CH 2 O - R - OH OH
O - CH 2 - CH - COOH O - CH 2 - CH - CH 2 OH
Фосфатидилсерин Фосфатидилглицерин
CH 2 O - C - R O CH 2 O - C - R
| || R /// - C - OH 2 C ||
CHO - C - R 1 O | CHO - C - R 1
| || R // - C - ONS ||
CH 2 O - R - OH O | CH 2 O - R - OH
O - CH 2 - CH - CH 2 O - R - OH 2 C O
|
|
Дифосфатидилглицерол Фосфатидилинозитол
(кардиолипиден)
Съставлява до 15% от всички миокардни липиди.Понякога във фосфоацилглицеролите една от мастните киселини се заменя с алдехид, при което се образуват плазмалогени.
CH 2 - O - CH \u003d CH - R /
CH - O - C - R 2
CH 2 - O - R - OH
В тези структури ясно се вижда, че съединенията имат две различни области: неполярна хидрофобна „опашка“ и полярна хидрофилна „глава“. Такава двойствена природа се нарича алифатична (от гръцки - двустранно и чувство).
2.Сфингомиелини (фосфосвинголипиди).
Те се състоят от една молекула мастни киселини, холин, фосфорна киселина и суингозинов алкохол. Сфингозините са семейство аминоалкохоли с дълга ненаситена верига, които се различават по дължина на веригата. FA се прикрепя към аминогрупата на сфингозин чрез амидна връзка и се образува керамид. Добавянето на фосфорилхолинов остатък завършва образуването на суингомиелин:
CH 3 - (CH 2) 12 - CH \u003d CH - CH - CH - CH 2 O - R - O - CH 2 - CH 2 - N + (CH 3) 3
Сфингомиелини
Сфингомиелините също имат амфипатични свойства. Сфингомиелините се намират в значителни количества в клетъчните мембрани на нервната и мозъчната тъкан.
3. Гликолипиди (гликосфинголипиди).
Те също са на базата на керамид. Има цереброзиди и ганглиозиди. Друга последователна липидна биосинтетична реакция е ензимното превръщане на керамидните прекурсори в цереброзиди и ганглиозиди чрез свързване на въглехидратна група към керамида. Цереброзидите и ганглиозидите се намират главно (макар и не изключително) в клетъчните мембрани на нервната и мозъчната тъкан и също проявяват амфипатични свойства. Протозои цереброзидисъдържат монозахаридна група, свързана чрез гликозидна връзка към крайната -ОН група на церамида. По правило това е глюкоза или галактоза.
CH \u003d CH - (CH 2) 12 - CH 3
Около 25% от цереброзидите могат да бъдат представени от сулфонирани (на 3-та позиция) галактоцереброзиди. Други церебродиди съдържат по-дълги олигозахаридни групи (2-10 остатъка), съдържащи аминозахари, N-ацитилзахариди и фруктоза. Ганглиозидите също съдържат олигозахаридна група, но се характеризират с наличието на поне един остатък от сиалова киселина. Един ганглиозид се различава от друг (това се отнася и за цереброзидите) по структурата на олигозахаридния компонент. Остатъците от сиалови киселини в ганглиозидите съдържат от 1 до 4.
Функции на гликолипидите:
1. Придаване на твърдост на мембраните.
2. Образуване на антигенни химични маркери на клетката, определяне на кръвната група (според системата ABO).
3. Образуване на химични маркери на етапите на клетъчна диференциация.
4. Регулиране на нормалния клетъчен растеж и по този начин се свързва с трансформацията на нормалните клетки в туморни клетки.
5. Придаване на способността на клетките да реагират с някои биологично активни съединения, например с бактериални токсини (холера, тетанус), гликопротеинови хормони, интерферони и вируси, т.е. всъщност това е рецепторна функция. Всички тези процеси са от голямо биологично значение. Наред с гликолипидите, гликопротеините също участват в много от тези процеси. Това подчертава значението на олигозахаридите за процесите, протичащи на повърхността на мембраната.
6.участват в междуклетъчните взаимодействия.
III. Липидни производни.
Това е много разнородна група от съединения. Сходството се ограничава само от слабата разтворимост във вода. Най-важните членове на тази група са стероиди, каротеноиди и сродни витамини от липиден характер.
1. Стероиди. Те се намират във всички организми, където изпълняват различни функции. При хората те играят ролята на полови хормони, емулгиращи агенти при смилането на липидите и участват в транспорта на липидите през мембраните. В кръвната плазма те действат като противовъзпалителни средства и като регулатори на някои метаболитни процеси. Всички стероиди имат подобна структура, която се основава на перхидроциклопентанофенантрен. Разнообразието от стероидни структури се дължи на различната степен на ненаситеност и наличието на няколко групи в различни позиции на циклите.
 |
Перхидроциклопентанофенантрен
Наличието на въглеводородна странична верига (C 8 - C 10) в 17-та позиция и хидроксилна група в позиция 3 е характерно за голяма група стероиди, наречени стероли. Най-важният и често срещан от тях е холестеролът. Той влиза като структурен компонент в клетъчните мембрани, но концентрацията му в мембраните варира (0 - 40% от общото съдържание на липиди в мембраните). Холестеролът (поради съдържанието на консолидирана структура, която е по-малко гъвкава от разширената въглеводородна верига) придава на мембраната по-голяма твърдост (якост). Холестеролът също е основният метаболитен прекурсор на други важни стероиди, включително жлъчни киселини и полови хормони. В някои тъкани холестеролът е предшественик на витамин D. Половите стероидни хормони: мъжки - тестостерон (причинява всички полови белези) и мастни: естрадиол и прогестерон (основно отговорни за регулирането на менструалния цикъл).
CH - (CH 2) 3 - CH - (CH3) 2
Холестерол
За първи път е изолиран от камъни в жлъчката, откъдето идва и името (chole - жлъчка, steros - твърд).
2. Каротеноиди.Каротеноидите се разделят на две групи: каротини и ксантофили. И двете групи включват водонеразтворими пигменти, които са широко разпространени в природата, повечето от тях се намират в растения и водорасли. Каротините са чисто въглеводородни съединения, докато ксантофилите съдържат кислородни групи. Каротините са по-широко разпространени в природата. Най-често срещаният е бета-каротинът. Това е C 40 въглеводород със силно разклонена структура и ненаситени връзки, в двата края на въглеводородната верига има идентични заместени пръстени. Всички други каротеноиди по същество могат да се разглеждат като варианти на тази структура. По време на ензимното разцепване на бета-каротин се образуват симетрично две молекули витамин А. В ретината на окото редуцираната алкохолна форма на витамин А (ретинол) се превръща от ензим в окислена алдехидна форма (ретинал), която образува комплекс с различни протеини, свързващи ретината (опсини). Такъв комплекс е първичният светлинен рецептор във фоточувствителните клетки, които след това предават информация на клетките на нервната система. Има два вида фоточувствителни клетки:
1. Пръчици – възприемат слаба светлина и са нечувствителни към цвета.
2. Колбички – възприемат ярка светлина и отговарят за формирането на цветно изображение.
Оказа се, че в пръчките има само един оксин (родопсин). Шишарките съдържат поне 3 вида оксини. Те образуват комплекси с ретината, за да образуват рецептори, чувствителни към сини, червени и зелени цветове. Комбинацията от тези цветове дава различни нюанси. Общо човешкото око може да различи 256 000 нюанса. Установено е, че ретината се свързва ковалентно с опсина, образувайки комплекс родопсин-ретинал. В този случай се свързва цис-формата на ретината, която през много междинни етапи преминава в транс-изомера. В този случай една или повече междинни форми участват в генерирането на много малки електрически потенциали, които се активират нервна система. Цикълът на трансформации на фоточувствителния пигмент по време на зрителното възприятие завършва с регенерацията на цис-ретината.
Липидни витамини.
3. Витамин D. Необходим за нормалния метаболизъм на калция и фосфора, необходими за растежа на здрави кости и зъби. Недостигът му причинява рахит – заболяване, при което костите стават меки и пластични, което води до тяхната деформация. Витамин D се образува от прекурсор на стерол с UV лъчение. Един от важните прекурсори е 7-дихидрохолестеролът, който сам по себе си се получава ензимно от холестерола. От него се получава витамин D3. Има няколко форми на витамин D. Fora D 3 (холикалцеферол) присъства в млякото, маслото, черния дроб на рибите, които са основният екзогенен източник на този витамин. Витамин D3 може да се синтезира в кожата, при условие че се получава нормална порция слънчева светлина. Както ендогенният, така и екзогенният витамин D 3 се транспортира от плазмата до черния дроб и бъбреците, където се превръща в още по-активни хидроксилирани производни. Основната активна форма в организма е 1,25-дихидрокси производно.
4. Витамин Е.Основната структура на витамин Е (и има няколко форми) се нарича токоферол.Най-активната форма на витамин Е е алфа-токоферолът. При плъховете витамин Е е необходим за възпроизводството (това все още не е доказано за хората). При някои животни липсата на витамин Е води до мускулна дистрофия. Вит Е обикновено се добавя по време на обработката на храната, тъй като той, притежавайки антиоксидантни свойства, предотвратява развалянето на храната поради окисляване. В живите клетки витамин Е също действа като антиоксидант, заедно с витамин С и глутатион.
5. Витамин К.Недостигът му забавя съсирването на кръвта. Структурата му се основава на бицикличния нафтохинонсистема, към която е прикрепена дълга въглеводородна верига. То може да дойде от храна (пресни зеленчуци, сирене) или може да бъде стимулирано от бактерии в стомашно-чревния тракт. Неговият коагулационен ефект се определя от участието в синтеза на протромбин, който след това преминава в тромбин.
Биомембрани.
Биомембраните обхващат клетката и много органели в клетката. Това са много активни биологични системи, отговорни за такива процеси като селективен транспорт на вещества в и извън клетката и субклетъчните отделения (формации), свързването на хормони и други регулаторни молекули, реакции, катализирани от ензими, предаване на електрически импулси и дори синтеза на АТФ. Мембраните се различават една от друга и различните видове мембрани имат различни видове функционална активност. Например, само вътрешната митохондриална мембрана е отговорна за биосинтезата на АТФ.
Химичен състав:Мембраните са съставени от липидни и протеинови молекули. Относителното им количество се различава значително за различните мембрани, като варира от 20% протеин + 80% липиди до 75% протеин + 25% липиди. Въглехидратите под формата на гликопротеини и гликолипиди съставляват 0,5 - 10% от мембранното вещество.
липиден двуслой.Биомембранните липиди са подредени в два слоя. Всеки монослой е съставен от сложни липиди (и понякога холестерол), подредени по такъв начин, че неполярните хидрофобни опашки на молекулите са в близък контакт една с друга; в същия контакт - полярни хидрофилни елементи. Всички тези взаимодействия са изключително нековалентни. Два монослоя са комбинирани, ориентирани „опашка“ към „опашка“, така че се образува двуслойна структура, която има неполярна вътрешна част и две полярни повърхности. Дебелината на всеки двуслой е ≈ 3,5 - 4,0 nm. Липидният състав на монослоевете е различен. Например, цереброзидите и ганглиозидите обикновено се намират във външния монослой на клетъчната мембрана. Степента на тази разлика в липидния състав на монослоевете е различна за различните мембрани и може да се промени, тъй като клетката участва в различни процеси и с остаряването си. Липидните молекули могат да се преместват от един монослой в друг. Подвижността (твърдостта) и течливостта на липидния двоен слой се определя от вида и дължината на въглеводородните групи, които изграждат мастните киселини и сфингозините, както и от съдържанието на холестерол. Повишената твърдост се определя от повишено съотношение на наситени към ненаситени вериги и повишено съдържание на холестерол. Физическите свойства на мембраните също зависят от вида и разположението на протеините, свързани с липидния двоен слой.
Мембранни протеини . Цялостната структура на мембраната включва протеини, прикрепени към липидния двоен слой по един от двата начина:
1.свързани с хидрофилната повърхност на бислоя - (повърхностни мембранни протеини);
2. потопени в хидрофобната зона на липидния бислой - (интегрални мембранни протеини).
Всяка мембрана съдържа и двата вида протеини, но различните видове мембрани имат различни съотношения на тези протеини. Повърхностните протеини с техните полярни странични групи от аминокиселинни остатъци, разположени на повърхността на протеиновата молекула, са нековалентно свързани с хидрофилната повърхност на липидния двоен слой. Интегралните протеини се отличават със степента на потапяне в хидрофобната област на двуслоя. От двете страни на мембраната някои протеини не могат да проникнат много дълбоко в двойния слой, достигайки около половината от слоя, докато други протеини, така да се каже, пробиват двойния слой със своята структура. Протеините, подобно на липидите, могат да променят местоположението си в мембраната. Това движение може да бъде насочено по монослоя, може да бъде ротационно в монослоя или дори в двуслоя, може да бъде насочено от една повърхност на двуслоя към друга. Протеините могат да претърпят промени в тяхната конформация, което вероятно засяга конформацията на съседните протеини.
Функции на мембранните протеини : Всички мембранни протеини (както повърхностни, така и интегрални) могат да бъдат разделени на 2 големи групи въз основа на тяхната роля в състава на мембраните:
1. структурни протеини;
2. динамични протеини.
Структурните протеини спомагат за поддържането на структурата на цялата мембрана. Често протеините от този тип имат удължена форма и са разположени върху хидрофилната повърхност на липидния слой, действайки като молекулна лента. Динамичните протеини са протеини, които са пряко включени в клетъчните процеси, протичащи върху мембраната. Във всички типове клетки обикновено се разграничават три класа такива протеини:
1. транспортни протеини (участват в транспорта на съединения в и извън клетките;
2. каталитични протеини (играят ролята на ензими в реакциите, протичащи върху мембраната);
3. протеини - рецептори (специфично свързват определени съединения - хормони, токсини, невротрансмитери от външната страна на мембраната, което служи като сигнал за промяна на химичните процеси в мембраната или вътре в клетката).
мембранен транспорт.
Трансфер (транспорт) на съединения вътре или извън клетката, както и техният транспорт между цитоплазмата и различни субклетъчни органели (митохондрии, ядро и др.) Се определя от мембраните. Транспортните свойства на мембраната определят нейната полупропускливост - някои съединения могат да проникнат през нея, а други не. В зависимост от конкретното съединение и вида на мембраната се осъществява пасивен или активен транспорт.
1 .Пасивният транспорт е транспорт по концентрационния градиент без разход на енергия от клетката. Такъв транспорт понякога се нарича дифузия. Правете разлика между проста и улеснена дифузия.
Простата дифузия се осъществява без взаимодействие с мембранния протеин. Преносът на вещества през липидни двойни слоеве се осъществява през пори в мембраната, образувани от празнини в двуслойния слой, или през канали, образувани от мембранни протеини. Така се пренасят например вода, някои органични йони и някои съединения от липиден тип. Улеснената дифузия включва молекули-носители, обикновено мембранен протеин. Такъв процес обикновено започва със свързването на прехвърленото съединение с транспортния протеин. Транспортният протеин се доближава до противоположната страна на мембраната, освобождава това съединение, приема първоначалното си състояние и отново е готов да изпълнява функциите си. Друга версия на трансфера: протеинът не е в състояние да премине през двуслойния слой и самото свързано съединение преминава от един протеин в друг, докато не е от противоположната страна на мембраната и се освобождава там.
2 .Активен транспорт е случаят, когато преносът изисква разход на енергия от клетката. Използват се също АТФ енергия и други енергийни източници. В този случай също е необходимо участието на мембранни протеини и транспортната система може да се окаже мултимолекулна. Енергията може да се изразходва на всеки етап от прехвърлянето. В някои случаи един от етапите може да бъде стимулиран от хормон.
Една от най-важните системи - K + , Na + - помпа. Помпата е един протеин, наречен Na +, K + - ATPase. Молекулата на този протеин прониква през цялата мембрана и е олигомер с молекулно тегло 250 000. Механизмът на действие включва няколко етапа:
1. АТФ молекула от цитоплазмата се свързва с активния център на една от АТФазните субединици. Свързването на АТФ се придружава от свързването на 3 Na + от цитоплазмата.
2. Фосфорирането на АТФ-аза (поради АТФ) предизвиква конформационен преход в структурата на протеина, което води до преориентирането му по такъв начин, че свързаният Na + се екскретира през канал, отворен към външната страна на мембраната.
3. След отстраняването на Na +, 2K + отвън се прикрепват към йон-свързващите центрове на фосфоризирания протеин.
4. Отстраняването на фосфатната група чрез хидролиза предизвиква нов конформационен преход, възстановявайки първоначалната конформация, която освобождава 2K + в канала, сега отворен вътре в клетката.
По този начин, за всяка изразходвана ATP молекула, ATPase пренася 3Na + и 2K + (съответно от клетката и в клетката).
Мускулните клетки съдържат Ca ++ - ATPase помпа, която контролира мускулната контракция, а освобождаването на Ca ++ причинява мускулна контракция.
мембранни рецептори.Отговорът на живите клетки на експозиция различни видовебиологично активни вещества (хормони, различни токсини, невротрансмитери и др.) започва със свързването на веществото към външната повърхност на клетъчната мембрана на определено място, наречено рецептор. Обикновено това са протеини, но могат да бъдат и други вещества, като ганглиозиди. Помислете за работата на някои рецептори.