Лимфни възли - брой, структура, увеличение (лимфаденопатия). Лимфни синуси и клетъчна миграция
Страница 6 от 6
ЛИМФНИТЕ ВЪЗЛИ
Лимфните възли (hodi lymphatici) са най-многобройните органи имунна система, служат като биологични филтри за протичащата през тях лимфа (тъканна течност). Те лежат на пътеката лимфни съдовеот органи и тъкани до лимфни каналии лимфни стволове. Лимфните възли обикновено са разположени на групи. Може да има два или повече, а понякога и няколко десетки възли в група. Например, групата на горните мезентериални възли има 66-404, аксиларни - 12-45, повърхностни ингвинални - 4-20 възли.
всеки лимфен възел(nodus lymphaticus) е покрит отвън със съединителнотъканна капсула, от която вътре в органа се простират тънки клони - напречни греди, капсулни трабекули(трабекули). На мястото, където еферентните лимфни съдове напускат лимфния възел, възелът има лека депресия - порти(хилум). В областта на портата капсулата се удебелява доста силно, образувайки портално (хиларно) удебеляване, повече или по-малко дълбоко изпъкнало в възела. Порталните трабекули се простират от порталното удебеляване в паренхима на възела. Най-дългите от тях се свързват с капсулни трабекули.
В лимфните възли, към които тече лимфа от крайниците (ингвинални, аксиларни) и които също се наричат оматични, по-често една порта, във висцерални (мезентериални, трахеобронхиални) - до 3-4 порти. През портата артериите, нервите влизат в лимфния възел, вените и еферентните лимфни съдове излизат.
Вътре в лимфния възел, между трабекулите, е ретикуларната строма. Представлява се от ретикуларни влакна и ретикуларни клетки, които образуват триизмерна мрежа с бримки с различни размери и форми.
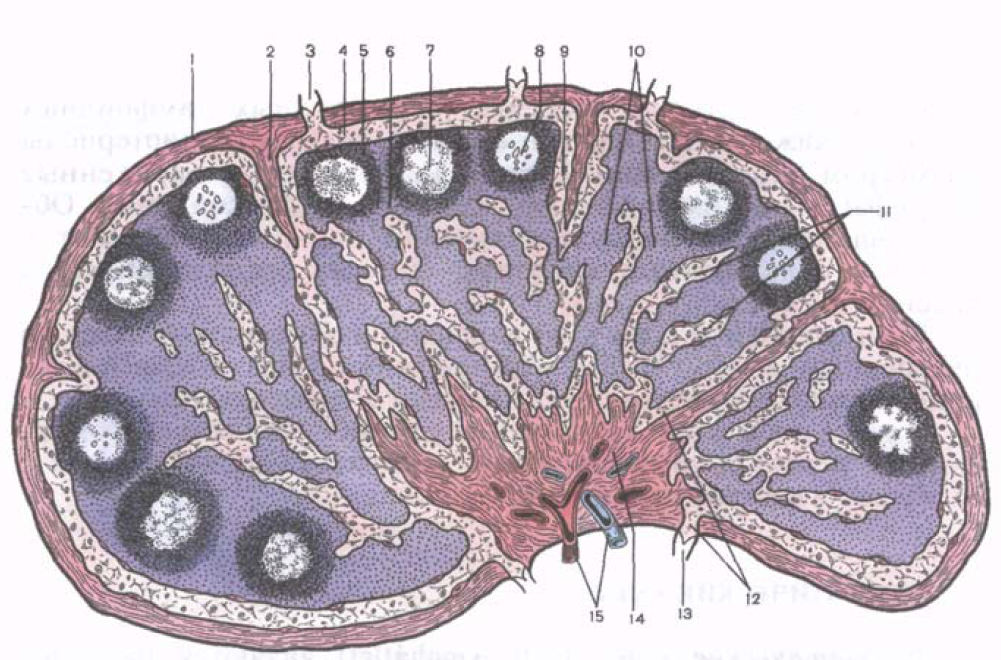
Ориз. 39. Микроскопска структуралимфен възел.
1 - капсула; 2 - трабекула; 3 - привеждане на лимфния съд; 4 - субкапсуларен лимфен синус; 5 - кортикално вещество; 6 - паракортикална (зависима от тимуса) зона; 7 - лимфоиден възел; 8 - центърът на възпроизвеждане на лимфоидния възел; 9 - кортикален лимфен синус; 10 - месести нишки; 11 - церебрални синуси; 12 - портален синус; 13 - еферентен лимфен съд; 14 - портално удебеляване; 15 - кръвоносни съдове.
Клетъчните елементи на лимфоидната тъкан са разположени в бримките на ретикуларната строма.
Паренхимът на лимфните възли е разделен на кортикален и медулен (фиг. 39). Кората е по-тъмна на оцветени хистологични срезове поради плътно разположени клетъчни елементи, по-близо до капсулата и заема периферните части на възела. Запалка медула(медула) лежи по-близо до вратата на възела и заема централната му част. В кортикалното вещество има заоблени образувания с диаметър 0,5-1,0 mm - лимфоидни възли(лимфоидни възли). Има лимфоидни възли без център за размножаване и с развъден център(зародишен център, centrum germinale).
Около лимфоидните възли е разположена дифузна лимфоидна тъкан. В него се разграничава кортикално плато, което включва области от лимфоидна тъкан между нодулите - интернодуларната зона. Съставът на кортикалното плато включва и тъкан, разположена извън лимфните възли, между тях и капсулата. Навътре от нодулите, директно на границата с медулата, се откроява ивица лимфоидна тъкан - перикортикалното вещество или тимус-зависима (паракортикална) зона(paracortex, s.zona thymodedependens), съдържащи предимно Т-лимфоцити. В тази зона има посткапилярни венули, облицовани с кубичен ендотел, през стените на които лимфоцитите мигрират в кръвния поток.
Паренхимът на медулата е представен от нишки от лимфоидна тъкан - меки кичури(chordae medullares). Те се простират от вътрешни отделикортикална субстанция към вратата на лимфния възел и заедно с лимфоидните възли образуват B-зависима зона. Нишките на целулозата са свързани една с друга, което води до сложни тъкани.
Паренхимът на лимфния възел е пронизан от гъста мрежа от тесни канали - лимфни синуси(sinus lymphatici), през който изтича постъпващата във възела лимфа субкапсуларен (маргинален) синус(sinus subcapsularis) към порталния синус. По протежение на капсулните трабекули лежат кортикални синуси(sinus corticales) и медула(синус медуларес). Последните достигат до вратата на лимфния възел (портално удебеляване) и се вливат в разположения тук портален синус. В лумена на синусите има малка бримкова мрежа, образувана от ретикуларни влакна и клетки, в чиито бримки могат да се забият чужди частици, мъртви и туморни клетки.
Развитие и възрастови особеностилимфни възли.Лимфните възли и тяхната строма се развиват от мезенхима в близост до възникващите плексуси на кръвоносните и лимфните съдове, започвайки от 5-6-та седмица от живота на ембриона. Отметките на лимфните възли в различни области на човешкото тяло се образуват в различни периодипреди раждането и дори след него.
В процеса на развитие на възела луменът на лимфния съд се превръща в субкапсуларен (маргинален) синус. Междинните синуси се развиват на базата на разклонен лимфен плексус, между съдовете на който растат нишки от ембрионална съединителна тъкан. Тук се вливат клетките от лимфоидната серия. Започвайки от 19-та седмица, в някои лимфни възли може да се види очертаната граница между кората и медулата. Лимфоидните възли в лимфните възли започват да се образуват още в пренаталния период. Центровете за възпроизвеждане в лимфоидните възли се появяват малко преди и малко след раждането. Основните свързани с възрастта процеси на формиране в лимфните възли завършват до 10-12 години.
Свързани с възрастта промени в лимфни възли(намаляване на количеството лимфоидна и пролиферация на мастна тъкан) се наблюдават още в юношеска възраст. Съединителната тъкан расте в стромата и паренхима на възлите, появяват се групи от мастни клетки. В същото време броят на лимфните възли в регионалните групи намалява. Много малки лимфни възли са напълно заменени от съединителна тъкан и престават да съществуват като органи на имунната система. Лежащите близки лимфни възли, често със среден размер, се сливат един с друг и образуват по-големи възли със сегментна или лентова форма.
Съдове и нерви на лимфните възли.Всеки лимфен възел включва от 1-2 до 10 малки артериални клона от най-близките артерии. Венулите, образувани от капилярите, се сливат във вени, отиват до портите на органа и напускат възела заедно с еферентните лимфни съдове.
Вегетативна нервни влакналимфните възли се получават от плексусите, разположени до артериите, както и от нервните стволове, преминаващи близо до възлите.
Въпроси за самоповторение и самоконтрол
1. Кои органи са централните органи на имунната система, кои са периферните и защо?
2. Какви морфологични особености са характерни за централните органи на имунната система и кои за периферните?
3. Спомнете си от какви структурни елементи се състоят костният мозък и тимусът?
4. Какво е общото в структурата на периферните органи на имунната система?
5. В стените на кои органи се намират лимфоидни възли? Каква е тяхната структура?
6. Какви структурни елементи на лимфоидната тъкан са включени в бялата пулпа на далака?
7. Какви образувания има в лимфните възли, през които лимфата тече от аферентните лимфни съдове към еферентните?
Държава Челябинск медицинска академия
Катедра по хистология, цитология и ембриология
Лекция
Периферни органи на хемопоезата и имуногенезата
Списък със слайдове.
1. Ретикуларна тъкан на лимфния възел (335).
2. Ретикуларна тъкан на далака (схема) (336).
3. Лимфен възел (схема) (339).
4. Лимфен възел. Месести корди (диаграма) (338).
5. Лимфен възел. Синусите (340).
6. Далак (диаграма) (341).
7. Далак (342).
8. Далак. лимфоиден фоликул. централна артерия (343)
9. Церебрални синуси на нестимулиран лимфен възел (390).
10. Церебрални синуси на стимулирания лимфен възел (391).
11. Ретикуларни влакна (схема) (337).
12. Плазматични клетки на миши далак (373).
13. Макрофаг, лимфоцити, бластоцити в лимфен възел (371).
14. дендритна клетка и лимфоцит (376).
15. Лимфобласт и лимфоцити в далака (369).
16. Лимфоидни фоликули на плака на Peyre (355).
17. B-зона на лимфен възел на имунизирана мишка със силна бластна трансформация и активни посткапилярни венули (386).
Планирайте.
1. основни характеристикипериферни органи на хемопоезата и имунната защита.
2. Хистофизиология на лимфните възли: източник на развитие, структура, характеристики на кората и медулата, Т- и В-зони, функции.
3. Характеристики на лимфоидната тъкан.
4. Хистофизиология на далака: източник на развитие, структура, характеристики на кората и медулата, Т- и В-зони, функции.
5. Т и В лимфопоеза.
Цел на лекцията:
1. Запознаване с морфологичните и функционални характеристики на периферните органи на хематопоезата и имунната защита.
2. Дайте понятието лимфоидна тъкан.
3. Опишете подробно характеристиките на Т и В лимфопоезата.
Периферните хемопоетични органи включват лимфни възли, далак, сливици, апендикуларен процес и лимфоидни фоликули в стената храносмилателен тракт, дихателни пътищаи пикочната система. В периферните органи на хематопоезата има среща на имунокомпетентни клетки с антигени. След това се активират имунни отговори, които се основават на антиген-зависима диференциация на лимфоцитите, което води до образуването на ефекторни клетки, които инактивират антигена, включително цитотоксични Т-убийци и плазмени клетки, които произвеждат антитела.
Лимфните възли.
При възрастен човек има до 1000 лимфни възли, чийто размер варира от главичка на карфица до малко бобено зърно (средно 1 см).
Лимфният възел има бобовидна форма и е разположен по хода на лимфните съдове. От конвексната повърхност в нея навлизат аферентните лимфни съдове. Вдлъбнатата повърхност на лимфния възел се нарича порта. В областта на портата артерия и нерви влизат в лимфния възел, а вената и лимфният съд излизат. От повърхността лимфният възел е покрит с капсула, състояща се от плътна съединителна тъкан, в чиито дълбоки слоеве има гладкомускулни клетки, които насърчават движението на лимфата. Съдържанието на гладкомускулни клетки и еластични влакна в капсулите на лимфните възли не е еднакво. По този начин ингвиналните и мезентериалните лимфни възли при хората са богати на мускулни клетки. В същото време сенилните лимфни възли се характеризират като правило с повишено съдържание на еластични влакна. Около съединителнотъканната капсула лежи мощен слой мастна тъкан, която постоянно обгражда лимфния възел.
Влакнестите трабекули се простират от вътрешната повърхност на капсулата и анастомозират една с друга в централните части на лимфните възли. Стромата на лимфните възли е представена от свободна неоформена съединителна тъкан и ретикуларна тъкан. В ретикуларната тъкан на лимфните възли се появяват клъстери от лимфоцити. Ретикуларната тъкан, инфилтрирана с лимфоцити, се нарича лимфоидна тъкан. От капсулата на лимфния възел лимфоидната тъкан образува сферични клъстери, които се наричат вторични възли или лимфоидни фоликули. От лимфоидните фоликули нишките се простират във вътрешността на възела, наречени медуларни връзки (мозъчни връзки). Белодробните връзки многократно анастомозират един с друг. Наличието на вторични възли (лимфоидни фоликули), разположени по периферията, и пулпидни връзки (мозъчни връзки), заемащи централната част на възела, позволяват да се изолират кортикалната и медулата на лимфния възел. Между кортикалната и медулата се намира паракортикалната зона.
Кортикалното вещество е разположено по периферията на възела и е представено от лимфоидни фоликули, в които централната част е леко оцветена, а периферната част е по-тъмна. Централната част се нарича светъл център, който съдържа предимно големи (незрели) лимфоцити. Поради факта, че в тази зона се намират множество митози, тя се нарича център на възпроизвеждане. Поради факта, че под въздействието на антигена в тази зона процесите на пролиферация на лимфоцитите протичат бързо и доста интензивно, тя се нарича реактивен център. Светлинните центрове (възпроизводствените центрове) не се намират във всички фоликули и размерите им са много променливи. Така че при животни, държани в асептични условия, няма светлинни центрове. Периферната, тъмно оцветена част на лимфоидния фоликул се нарича зона на мантията, в която преобладават малките лимфоцити. Лимфоидните фоликули не са постоянни образувания: те могат да се появяват и изчезват отново.
Лимфоидните фоликули и пулпичните връзки са заобиколени от синуси на лимфни възли, които са подобни на прорези пространства, пълни с ретикуларна тъкан. Лимфата тече през синусите. Има няколко вида синуси, включително маргинален синус, междинен кортикален синус, церебрален синус и портален синус. Маргиналният синус (субкапсуларен) е ограничен от съединителнотъканната капсула на лимфния възел и повърхността на лимфоидния фоликул. Междинният кортикален синус е ограничен от повърхността на трабекулите и страничната повърхност на лимфоидния фоликул. Мозъчните синуси лежат между разклонените месести връзки. Порталният синус (терминален синус) се намира в областта на портата на лимфния възел. Стената на всички синуси на лимфния възел е облицована със специални, модифицирани ретикуларни клетки - брегови клетки. Крайбрежните клетки придобиват характеристики на ендотелни клетки с висока фагоцитна активност. Между крайбрежните клетки има многобройни пори, които създават широки връзки между лумените на синусите и лимфоидната тъкан на пулпичните връзки и лимфоидните фоликули. Благодарение на крайбрежните клетки, лимфата, протичаща през синусите, се филтрира и изчиства от чужди съединения.
В лимфните възли се разграничават Т- и В-зони. B-зоната е представена от лимфоидни фоликули и месести връзки. В тази зона антиген-зависимата диференциация на В-лимфоцитите се извършва под въздействието на специфична микросреда, която включва ретикуларни клетки, дендритни клетки от първи тип, макрофаги и малък брой Т-лимфоцити. Т-зоната е представена от лимфоидната тъкан на паракортикалната зона. В тази зона протича антиген-зависима диференциация на Т-лимфоцитите под въздействието на специфична микросреда, която включва ретикуларни клетки, дендритни клетки от втори тип (интердигитиращи клетки), макрофаги, както и малък брой В-лимфоцити и плазмени клетки. .
В паракортикалната зона има специализирани участъци на кръвния поток - "посткапилярни венули", чиято стена е облицована с кубични или призматични ендотелни клетки. Тези ендотелни клетки имат добре развит ендоплазмен ретикулум, цитоплазмени везикули и микровили. Посткапилярните венули са мястото на инвазия на Т и В лимфоцитите в лимфните възли. Т и В лимфоцитите първоначално се прилепват към ендотелните клетки и след това преминават между тях, придобивайки цитоплазмена полярност. След проникване в лимфоидната тъкан, лимфоцитите заселват Т и В зоните, където се извършва тяхната антиген-зависима диференциация.
Лимфните възли са богато инервирани. Амиелинизираните и немиелинизираните нервни влакна навлизат в лимфния възел заедно с артерията.
Функции на лимфните възли.
1. Лимфопоетична (хемопоетична) функция.Лимфата, протичаща през синусите на лимфния възел, се обогатява със зрели Т и В лимфоцити, напускайки лимфоидната тъкан през порите между крайбрежните клетки.
2. Имунологична функция. Благодарение на Т и В лимфоцитите, образувани в лимфните възли, последните участват в регулацията на клетъчния и хуморален имунитет.
3. Бариерна (защитна) функция.Лимфата, протичаща през синусите, се изчиства от чужди съединения поради фагоцитната активност на крайбрежните клетки.
4. Депозираща функция.Обикновено определено количество лимфа се задържа в лимфния възел и се изключва от лимфния поток. Ако е необходимо, той отново навлиза в лимфната циркулация.
5. Обменна функция.Лимфните възли вземат Активно участиев метаболизма, включително протеини, мазнини, въглехидрати и други съединения. Хранителните вещества в лимфата могат да бъдат уловени от макрофагите и да бъдат разградени от лизозомни ензими.
развитие. Лимфните възли се развиват на 8-10-та седмица от ембриогенезата от натрупвания на мезенхимни клетки в близост до кръвоносните съдове и по протежение на лимфните съдове. По периферията на тези натрупвания от мезенхима се образува съединителнотъканна капсула. Между мезенхима на рудимента на възела и капсулата се образува пространство - субкапсуларен синус, от който се отклоняват кортикалните перинодални синуси, разделящи възлите един от друг; церебралните синуси се отклоняват от перинодуларните синуси. Трабекулите се простират от капсулата на съединителната тъкан дълбоко в мезенхимния рудимент на възела. Мезенхимните клетки на рудимента на лимфните възли се диференцират в неговата ретикуларна строма. Скоро стволовите клетки нахлуват в зачатъците на лимфните възли и започва миелопоезата, която продължава за относително кратък период от време. На 16-та седмица В-лимфоцитите нахлуват в центъра на лимфните възли, няколко дни по-късно те (В-лимфоцитите) нахлуват и в периферните им части, а последни нахлуват Т-лимфоцитите. От този момент започва лимфопоезата в лимфните възли, която продължава преди и след раждането. До 20-та седмица от ембриогенезата лимфните възли придобиват чертите на крайните (окончателни) лимфни възли.
Структура. Лимфните възли са разположени по протежение на кръвоносните и лимфните съдове, имат овална или бобовидна форма. Отвън лимфните възли са покрити със съединителнотъканна капсула, от която се простират трабекули. Капсулата и трабекулите включват колагенови и еластични влакна и гладки миоцити. На вдлъбнатата повърхност на лимфните възли има порти. Артерията и нервите влизат в портата, вените и еферентният лимфен съд излизат. Аферентните лимфни съдове влизат от противоположната изпъкнала страна. Стромата на лимфните възли е ретикуларна тъкан, състоящ се от ретикуларни клетки и ретикуларни влакна. Диаметърът на лимфните възли е от 0,5 до 1 cm.
По периферията на лимфните възли има по-тъмна кортикална субстанция, представена от лимфни възли, а в центъра - по-светла медула, представена от медуларни връзки. Лимфните възли имат кортикална зона (кортекс), медула и паракортикална зона, разположени между кортикалната зона (лимфни възли) и нишките на медулата.
кортикална зона представени от лимфни възли (nodulus lymphaticus), чийто диаметър варира от 0,5 до 1 mm. Стромата на лимфните възли е представена от ретикуларна тъкан, предимно кръгово разположени ретикуларни влакна. Лимфните възли съдържат свободни макрофаги, дендритни клетки, В-лимфоцити и В-лимфобласти. В центъра на лимфните възли е светъл цвятцентър (centrum lucidum). Този център се нарича също зародишен център, тъй като В-лимфобластите се размножават тук. както и реактивния център, защото тук протича реакцията между макрофагите и антигените. С други думи, в светлинните центрове има активиране на лимфоцити от антигени, възпроизвеждане на лимфобласти (имунобласти, плазмобласти) и фагоцитоза на антигени и лимфоидни клетки.
Макрофаги лимфните възли изпълняват фагоцитна функция и преработват антигени от корпускуларно състояние в молекулярно състояние, натрупвайки антигенни молекули до такова количество, което може да причини антиген-зависима диференциация на В-лимфоцитите.
Дендритни клетки лимфните възли са макрофаги, които са загубили способността си да фагоцитират. В дендритните клетки има процеси, слабо развити органели от общо значение. Цитоплазмата на тези клетки е слабо оцветена, на повърхността им има рецептори за имуноглобулини. Имуноглобулините са прикрепени към тези рецептори, а антигените са прикрепени към свободните краища на имуноглобулините. Антигените на дендритните клетки, заедно с антигените на макрофагите и с участието на Т-хелперите, стимулират В-лимфоцитите към пролиферация, диференциация и функционална активност (производство на антитела).
Откъде идват В-лимфоцитите в кортикалната зона? Те идват тук от червения костен мозък с кръвен поток. В-лимфоцитите, навлизащи в лимфните възли, са изложени на антигени, фагоцитирани от макрофаги, антигени, задържани върху рецепторите на дендритни клетки, и лимфокини, секретирани от Т-хелперите. Под въздействието на всички тези влияния В-лимфоцитите претърпяват бластна трансформация, пролиферация и антиген-зависима диференциация. В резултат на тази диференциация В-лимфоцитите се превръщат в ефекторни клетки - плазмоцити и клетки на паметта. Плазмените клетки са способни да произвеждат антитела, насочени към унищожаване на тези антигени, под влиянието на които са претърпели антиген-зависима диференциация. След това плазмените клетки и клетките на паметта навлизат в общия кръвен поток през посткапилярните венули и след кратка циркулация преминават в съединителната тъкан. В съединителната тъкан всеки от тях изпълнява своя собствена функция: плазмените клетки отделят антитела, а клетките на паметта, след като се срещнат с познат антиген, претърпяват диференциация на ефекторни клетки и влизат в имунна реакция, насочена към унищожаване на този антиген.
По този начин лимфните възли са зона на В-лимфоцити.Ако антиген влезе в тялото, тогава възлите се развиват на етапи.
Има 4 етапа на развитие. На етап I се образува светъл център; в този център лимфобластите претърпяват митотично делене.
В етап II този център се разширява; на участък от лимфен възел има до 10 митотични деления.
На етап III около светлите центрове на възлите се появява корона, състояща се от малки лимфоцити; броят на делящите се клетки намалява, светлият център се стеснява.
На етап IV делящите се клетки са единични; около тесен светлинен център има корона, състояща се главно от клетки на паметта.
Целият цикъл на промени от момента на навлизане на антигена до настъпването на етап IV (стадий на почивка) продължава 2-3 дни. Седмица след като антигенът влезе в тялото, мозъчните връзки се разширяват, броят на лимфоцитите и плазмоцитите в синусите на лимфните възли се увеличава. Ако антигените не попаднат в тялото (стерилни условия), тогава в лимфните възли няма светлинни центрове.
Паракортикална зона (паракортекс) се намира между лимфните възли и мозъчните връзки. Тази зона съдържа интердигитиращи клетки, Т-лимфоцити и Т-лимфобласти. Интердигитиращите клетки са наречени така, защото имат процеси, които проникват между краищата на процесите на съседни интердигитиращи клетки. Интердигитализиращите клетки са макрофаги, които са загубили способността си за фагоцитоза, съдържат слабо развити органели от общо значение и имат слабо оцветена цитоплазма. Интердигитиращите клетки произвеждат гликопротеини, които стимулират диференциацията на Т-лимфоцитите, а гликопротеините, разположени под плазмената мембрана, действат като рецептори, които задържат антигени, които участват в диференциацията на Т-лимфоцитите. В тази зона има кооперативно взаимодействие на имунокомпетентни клетки. При отстраняване на тимуса (тимектомия) паракортикалната (зависима от тимуса) зона е слабо изразена.
По този начин паракортикалната зона е зона на Т-лимфоцитите, или тимус-зависима зона.
Медулата на лимфните възли по-лек, образуван в резултат на преплитане на мозъчните връзки. Стромата на медулата също е ретикуларна тъкан. Съставът на мозъчните връзки включва плазмени клетки. B-лимфоцитите, макрофагите и ретикуларните клетки, т.е. мозъчните връзки са зоната на B-лимфоцитите. Кръвоносните капиляри протичат в мозъчните връзки.
Синусите на лимфните възли.Между капсулата и лимфните възли има субкапсуларен синус (sinus subcapsularis), между трабекулите и лимфните възли има перинодуларни синуси (sinus perinodularis), излизащи от субкапсуларния синус, между трабекулите и мозъчните връзки има церебрални синуси (синус medullaris), излизащ от перинодуларните синуси.
Синусите са облицовани с ретикулоендотелни клетки, тоест специални ендотелни клетки, подобни на ретикулоцитите. Сред ретикулоендотелните клетки има брегови клетки - макрофаги. Тези макрофаги фагоцитират антигени, намиращи се в лимфата, протичаща в синусите. Ретикулоендотелните клетки на синусите на повърхността на стената, съседна на капсулата и трабекулите, лежат върху базалната мембрана, а на повърхността, обърната към лимфните възли и мозъчните връзки, те лежат върху мрежата от ретикуларни влакна, които оплитат лимфните възли и мозъчни връзки. Това допринася за потока на лимфоцити и плазмени клетки от възлите и мозъчните връзки в лумена на синусите.
Функции на лимфните възли .
Хематопоетичната функция, която се състои в антиген-зависимата диференциация на лимфоцитите, което води до образуването на ефекторни клетки и клетки на паметта, участващи в имунните реакции, е функция на имунната защита.
Защитна функция, която се състои в това, че макрофагите фагоцитират бактерии, клетъчни фрагменти, антигени, които се намират в лимфата, протичаща през синусите.
Освен това лимфата се обогатява с лимфоцити. Лимфата се отлага в лимфните възли.
Лимфните възли на мезентериума на червата участват в метаболизма на липидите, които се абсорбират лимфни капиляривили и се транспортират до лимфните възли.
Лимфните възлиили лимфни възлиса органи на човешката лимфна система. Те са овални, кръгли, понякога с лентовидна форма и са разпространени по цялото човешко тяло, включително подмишниците, и са свързани помежду си с лимфни съдове. Размерите им варират от 0,5 до 50 мм, боядисани в розов цвят. Лимфните възли са мястото на концентрация на В, Т и други имунни клетки.
Лимфните възли играят ролята на капани и филтри за чужди частици, те са необходими за нормално функциониранечовешката имунна система. В лимфните възли има плътна концентрация на бели кръвни клетки (лимфоцити и макрофаги).
Лимфните възли са от клинично значение. Те се възпаляват или уголемяват различни заболяваниякакто инфекциозни, така и онкологични. Значението на състоянието на лимфните възли за определяне на наличието на рак е важно.
Лимфните възли не са част от човешката система за борба с болести, свързани с отравяне. токсични вещества, тези задачи се изпълняват от бъбреците и черния дроб.
Възпалените лимфни възли се диагностицират с биопсия. Болестите, които причиняват възпаление на лимфните възли, имат определен курс и локализация.
Структурата на лимфните възли
Лимфните възли са заобиколени от фиброзни капсули, а вътре фиброзната капсула се простира, за да образува трабекула. Субстанцията на лимфните възли е разделена на кортикална субстанция и медула, заобиколена навсякъде от кората, с изключение на хилуса, където медулата контактува с повърхността.
Тънките ретикуларни влакна, еластинът и ретикуларните влакна образуват поддържаща мрежеста структура - ретикуларна мрежа в рамките на възела, в която белите кръвни клетки са плътно концентрирани като фоликули в кората. На други места има само отделни левкоцити. Функцията на ретикуларната мрежа е не само да осигурява опора, нейната повърхност служи за адхезия на дендритни клетки, макрофаги и лимфоцити. Той насърчава метаболизма с кръвта през ендотелните венули и също така осигурява процесите, необходими за появата и узряването на имунните клетки.
Лимфният възел има канал - лимфен синус, облицован с ендотелни и ретикуларни клетки, фибробласти, който осигурява плавен поток на лимфата. По този начин субкапсуларният синус е синус, разположен директно в капсулата и неговият ендотел е продължен от аферентен лимфен съд. Лимфните синуси, заедно с паренхима, са единна морфологична и физиологична единица. От изпъкналата страна в капсулата влизат няколко аферентни съда, които отвеждат лимфата в синуса, разположен под капсулата. Този синус се нарича маргинален или циркулярен. Този синус е обширна формация, подобна на прорез, която покрива целия паренхим на възела, който се намира под капсулата и се прекъсва само в точките на проникване на трабекулите в паренхима. Точно там маргиналният синус преминава в интерстициалните (междинни) синуси, които по-късно са церебралните синуси, преминавайки в мозъчната част на възела, където по-нататъшната им връзка вече не се наблюдава. За разлика от предишните, церебралните синуси са големи и структурата им изглежда гъбеста поради факта, че техните ендотелни клетки са по-високи. Междинните и маргиналните синуси са в тясна връзка с лимфните фоликули, създавайки функционални връзки. Мозъчните синуси се сливат в портите на лимфните възли, като по този начин образуват еферентни съдове, които извеждат лимфата.
Стените на лимфните синуси са изградени от ендотелни клетки, които се наричат литориални. Някои от тези клетки са с форма на звезда и са оборудвани с израстъци, които пресичат синуса напречно и свързват противоположните му стени като мостове. Натрупването на тези клетки е филтър за протичащата през тях лимфа.
Разликата между лимфните синуси и кръвоносните съдове е, че те имат базална мембрана от страната на трабекулите и капсулата, докато няма такава мембрана от страната на паренхима на лимфния съд. Това може да се види, когато се гледа с електронен микроскоп.
Там, където има лимфна тъкан, ендотелните клетки са плътно притиснати към ретикуларните влакна, които заместват базалната мембрана. Цитоплазмата на ендотелните клетки на синусите се състои от тънък гладък везикуларен ендоплазмен ретикулум, а клетките, разположени в близост до лимфния паренхим, се състоят от лизозоми и гранулиран ендоплазмен ретикулум.
Местоположение на лимфните възли
Лимфните възли са разположени по дължината на лимфните съдове в групи от до 10 възли, както и в близост до кръвоносни съдове и големи вени.
Лимфните възли са разположени на групи, така че да създават бариера за разпространението на инфекции и ракови клетки. Те се намират например на следните места: коленни гънки, ингвинална област, лакътни гънки, подмишници. Лимфните възли, които се намират на шията, предпазват органите на главата и околошийните органи от инфекции и тумори.
Увеличени лимфни възли
Увеличаването на лимфните възли показва наличието на заболяване в областта, където се намира възелът. Най-често това се свързва с инфекциозни заболявания, и рядко с тумори.
Остър лимфаденит - възпаление на лимфен възел възниква, когато има гноен процес. Основните симптоми са увеличаване на лимфните възли, болка при усещане. По време на възпалителния процес върху възпаления лимфен възел кожата може да се зачерви. Може да се наложи хирургично отваряне, ако не се направи навреме, лимфният възел може да се спука и съдържанието му да проникне в околните тъкани - развива се флегмон.
При деца с туберкулоза се наблюдава увеличение на лимфните възли, което е един от симптомите на заболяването.
Също така, увеличаването на лимфните възли може да бъде свързано със заболяване като " котешка драскотина". В който микроб прониква през драскотина - Bartonella, която се пренася от котки.
Също така, увеличаването на лимфните възли е свързано с респираторни заболявания и е следствие от прекомерна реакция на имунната система към инфекция.
При сифилис увеличаването на лимфните възли в слабините се появява след появата на язва на гениталиите.
Продължителното увеличение на няколко лимфни възли може да показва наличието на ХИВ, бруцелоза, мононуклеоза, листериоза.
При тумори увеличаването на лимфните възли може да бъде свързано с разпространението на метастази, както и с развитието на тумор директно в лимфния възел (лимфогрануломатоза, лимфосаркома).
Паренхимът на лимфните възли е пронизан от гъста мрежа от канали (прорези) - лимфни синуси, през които лимфата, влизаща в възела, тече от субкапсулния (маргинален) синус към порталния. Непосредствено под капсулата на възела, между него и паренхима, има субкапсуларен (маргинален) синус, в който се вливат аферентните лимфни съдове. От субкапсулния синус до паренхима на възела, по протежение на капсулните трабекули, междинните синуси на кората и медулата напускат. Последните достигат до вратата на лимфния възел (хиларно удебеляване) и се вливат в порталния синус, от който произхождат еферентните лимфни съдове. Субкапсулният (маргинален) синус също се влива в порталния синус, покривайки паренхима на органа по периферията (под капсулата) и завършвайки в областта на портата на възела. Междинните синуси на кортикалното вещество преминават дълбоко в органа между лимфоидните възли, понякога се огъват, покривайки възела от едната или другата страна (кръгло-нодуларен кортикален синус). Разположени по протежение на капсулните трабекули, кортикалните синуси, от една страна, са в съседство със съединителната тъкан на трабекулите, а от друга страна, с лимфоидния паренхим на кортикалното вещество.
В медулата синусите са разположени по два начина.
Някои междинни церебрални синуси лежат между съседни месести връзки (интерпулмонални синуси). Други са (на хистологичен разрез) между пулпизните връзки от едната страна и хиларната трабекула от другата (трабекуларни синуси). Синусите на медулата са по-широки от субкапсуларните и кортикалните синуси. Тънките стени на синусите от страната на техния лумен са облицовани с удебелени ендотелни (крайбрежни) клетки. Маргиналният синус от страната на капсулата, както и стените на кортикалните, церебралните и порталните синуси, съседни на трабекулите и порталното удебеляване, имат непрекъснат слой крайбрежни клетки. Стените на синусите, ориентирани към лимфоидния паренхим, са изградени така, че през тях от кортикалната и медулата в лимфата и в обратна посокалимфоцити, макрофаги и други активно движещи се клетки могат лесно да проникнат. В лумена на синусите има фино петлична мрежа, образувана от ретикуларни влакна и клетки. В бримките на тази мрежа могат да се задържат чужди частици, микробни тела и туморни клетки, навлизащи в лимфния възел заедно с лимфата.
С помощта на сканираща микроскопия на корозивни и нативни препарати на V.
К. Шишло и А. А. Миронов (1990) показаха, че мрежата от ретикуларни клетки и техните процеси, както и влакна, които съществуват в синусите, служат за създаване на турбулентност в лимфния поток. Това улеснява филтрирането на лимфата през мрежата вътре в синусите. По отношение на клетките, покриващи стените на синусите, има две мнения. Редица автори разглеждат тези клетки като клетки, близки до ендотелните, като че ли са продължение на ендотела на аферентните лимфни съдове. Според друга гледна точка тези клетки са модифицирани ретикуларни клетки със способност за фагоцитоза. Името "крайбрежни" е дадено на тези клетки от N. Siegmund (1923). Y. Mori и K. Lennert (1969) наричат синусовите клетки ретотелни. Във фината структура на ретотелните клетки се забелязват разлики в зависимост от тяхната локализация - близо до маргиналния синус, междинните синуси на кората и медулата. По-специално, L. V. Chernyshenko (1966) посочва, че ендотелът на маргиналния синус на париеталните лимфни възли на коремната кухина при фетуси и новородени има клетъчна структура, докато в горните и предните панкреатични и горните стомашни възли има „синцитиална“ ” структура.
Повечето крайбрежни (ретотелни) клетки съдържат тънки нишки по дългата ос на клетките. Съседните клетки са свързани чрез десмозоми. За разлика от клетките на маргиналния синус, крайбрежните клетки на междинните синуси съдържат голям брой органели. Те съдържат множество везикули от гладкия цитоплазмен ретикулум и дълги тубули от гранулирания цитоплазмен ретикулум.
Както е показано от Ю. И. Бородин и В. Н. Григориев (1983), клетките, които образуват външната стена на синуса, са свързани помежду си чрез прилепване една към друга и образуване на сложни контакти като интердигитални. В същото време по протежение на контактите се появяват петна и зони на заличаване, което показва плътността на тези контакти. Формата на клетъчните ядра на външната стена на маргиналния синус е овална или удължена, а понякога дори пръчковидна. Хроматинът е финозърнест, разпределен главно по периферията на ядрото. Цитоплазмената мембрана образува гънки, обърнати както към лумена на синусите, така и към съседната съединителна тъкан. Понякога лимфоцитите "прилягат" много близо до клетките на външната обвивка на синуса и техните израстъци навлизат в инвагинациите на клетките на стената на синуса. Органелите са равномерно разпределени в цитоплазмата. Митохондриите най-често имат кръгла или овална форма, с малък брой кристи. Зърнестите и негранулираните цитоплазмени ретикулуми са слабо развити, преобладава грануларният ретикулум. Има голям брой свободно разположени рибозоми и политики под формата на вериги или розетки. Лизозомите са малко, електронно плътни, заоблени, с гранулирано съдържание. В перинуклеарната зона на цитоплазмата има ламеларен комплекс, състоящ се от сплескани цистерни и везикули. Везикулите са разположени в цялата цитоплазма на клетката, понякога се сливат, образувайки вакуоли. По този начин клетките, които образуват външната стена на синуса, наподобяват ултраструктурата на ендотела.
При учене вътрешна стенасинус В. Н. Григориев идентифицира 3 вида клетки, които се различават една от друга по морфологични особености. Клетките от 1-ви тип имат процеси и се свързват помежду си чрез просто кръстовище на процеси. В лумена между цитоплазмените процеси има снопове от ретикуларни влакна. Ядрата (рядко включени в разреза) заемат централната част на клетката. Формата им е овална, заоблена, понякога неправилна; хроматинът е финозърнест; органелите са разположени главно в зоната на перикариона. Митохондриите са кръгли или овални, с няколко кристи. Гранулираният цитоплазмен ретикулум е добре развит. Цитоплазмата на тези клетки, както и клетките, които образуват външната стена на синуса, съдържа голям брой свободни рибозоми и полизоми. Няколко лизозоми изглеждат като електронно-плътни заоблени образувания с гранулирано съдържание. Ламеларният комплекс е по-често разположен близо до ядрото. Пиноцитозната активност в тези клетки не се изразява. Вътрешната стена на маргиналния синус, представена от клетки от тип 1, има пори, в които понякога се виждат лимфоцити. Клетките от 2-ри тип, облицоващи вътрешната стена на синуса, за разлика от клетките от 1-ви тип, са сплескани, удължени. Те се свързват помежду си чрез просто напасване на процесите. Ядрата са овални или пръчковидни, хроматинът е финозърнест. Органелите на тези клетки, за разлика от органелите на клетки от тип 1, са слабо развити: видими са единични митохондрии, гладък цитоплазмен ретикулум и свободни рибозоми. Много пиноцитни везикули. Клетки от 3-ти тип се срещат рядко; те се намират директно в лумена на синусите в близост до клетките от 2-ри тип и са в тясна връзка с тях. Те имат овална форма, процесите на тези клетки образуват плътни контакти с тялото на собствената им клетка. Контурите на ядрената мембрана са извити, което показва увеличаване на повърхността на ядрото. Хроматинът е финозърнест. Органелите се развиват по същия начин, както в клетки от тип 1. Въпреки това, за разлика от последните, клетките тип 3 съдържат много лизозоми и пиноцитни везикули.
Размерът на крайбрежните клетки на церебралните синуси значително надвишава размера на крайбрежните клетки на вътрешната стена на маргиналния синус. Следователно, обемните съотношения на последните са значително по-големи от тези на крайбрежните клетки на церебралните синуси. С намаляване на функцията на имунната система при животни (и дори с имунодефицит), както G. Sainte-Marie и F.-S. Peng (1990), субкапсуларните синуси се разширяват. Изглеждат като че ли имат малко лимфоцити. В същото време синусите на медулата са пълни с множество макрофаги. Синусите се характеризират с проникване на клетки в нодуларната зона и това се случва на фона на атрофия на лимфоидните възли, наличието на така наречените тъмни лимфоцити, които се считат за увреждащи клетки. Според N. Chizini-Garcia, S. R. Machardo (1992), тук в синусите има и мастоцити, които се отличават с разнороден размер, форма и тинкториални свойства на гранулите. Възможно е те да съдържат хепарин, но той е по-малко в церебралните синуси, отколкото в останалата част на лимфните възли.
Във функционирането на лимфните възли, които пропускат лимфата през себе си и действат като биологични филтри, участват всички техни структури: кора и медула, множество лимфни синуси, разклонени в паренхима. В тази връзка интерес представляват данните за динамичните процеси в лимфните възли и ролята на Т- и В-лимфоцитите в тях. Според P. V. Pigarevsky (1991), нормално структурни компонентиТ- и В-зоните на висцералните и соматичните лимфни възли са 90-95% съставени от малки и средни лимфоцити и ретикуларни клетки и само 5% са клетъчни форми от плазмоцитната серия. В Т-зоната броят на Т-хелперите е по-голям от Т-супресорите, но това съотношение може да се промени през деня. Обикновено в рамките на Т- и В-зоните както на соматичните, така и на висцералните лимфни възли не настъпва преструктуриране поради образуването на клонинг на антиген-реагиращи клетки, необходими за развитието на имунен отговор. В Т-зоната рядко се наблюдават митотично делящи се клетки (от 0,02 до 0,06%, оказа се, че промени, свързани с възрасттатози показател не се влияе). А. Ю. Летягин (1991) смята, че дневната пространствено-времева структура на лимфните възли се състои от ритмично времеви морфофункционални комплекси. Избраният комплекс "А" се характеризира с увеличаване на броя на лимфоидните структури на лимфния възел в резултат на рециркулация и пролиферация на лимфоцити. Комплекс "В" се характеризира със значително и бързо намаляване на броя на лимфоцитите в органа поради емиграцията на тези клетки в периферното кръвообращение на фона на активирането на динамични фактори на микросредата и активността на дренажните системи, които осигуряват изпускане на течност от органа. Комплексът "B 2" се характеризира с нестабилен баланс на клетките и течността в органа. В същото време се наблюдава увеличаване на миграцията на лимфоцитите под активното влияние на факторите на микросредата. Всички тези данни са представени най-пълно в монографията на Ю. И. Бородин, В. А. Труфакин и др. (1992), постулирайки, че „процесът на миграция и рециркулация на лимфоидни елементи, дори при липса на имунизация, не е случаен. Лимфоцитите "знаят" границите на своята зона на действие, пътя на движение в лимфоидната система и периферните тъкани. Такъв ред възниква в процеса на онтогенезата както на отделен лимфоцит, така и на целия лимфоидна системаи корелира със способността за синтезиране и пренасяне на специфични рецептори.
I. T. Gegin и A. I. Krayushkin (1991) приемат лимфоидни възли с прилежащи територии, пулпидни връзки като продължение на възли дълбоко в медулата и привеждане на лимфните съдове като структурна и функционална единица в лимфните възли. В монографията на Ю. И. Бородин, М. Р. Сапин и др. (1992) обобщава отделни опити по някакъв начин да се раздели тъканта на лимфните възли на структурни единици. И така, G. Sainte-Marie et al. (1984, 1990), Ю. Е. Виренков, В. К. Шишло и Ю. Г. Антропова (1993) не са много убедително откроени като структурна единицаотделение за лимфен възел. Този сектор, който според авторите включва централната част на лимфния възел, съответстващ на паракортикалната област, няколко лимфоидни възли и пулпидни връзки, разположени в този сектор. централна частнаречена единица или Т-домейн. Последният се формира от стромални елементи, различни популации от Т-лимфоцити и малък брой В-лимфоцити, мигриращи към съответните им В-домени. Т-доменът се простира в екстранодуларното пространство, където се осъществява селективна миграция на лимфоцити през стените на венули с висок ендотел. Ю. Е. Виренков, В. К. Шишло и Ю. Г. Антропова (1993) показват, че всяко отделение е "посветено" само на един аферентен лимфен съд, 1-6 лимфоидни възли.
A. I. Krayushkin (1994) пише за наличието в лимфните възли на структурни и функционални части, съответстващи на органи и области на тялото, които са разнородни по функция и структура. Изследвайки лимфните възли на зайци от различни възрасти, той показа, че в централния мезентериален (висцерален) лимфен възел, който получава лимфа както от тънките, така и от дебелите черва, багрилата, въведени в стените на тези органи, при навлизане в лимфния възел, запълват синусите всеки в своята част.възел. Този автор също така отбелязва, че лимфните съдове от тънко червоподход от едната страна (отляво) на мезентериалния лимфен възел, а от дебелото черво - до противоположната (дясната) страна на този възел. Ю. И. Бородин (1994) също пише за съответствието на аферентните лимфни съдове с определена част от паренхима на лимфния възел. Според А. И. Краюшкин лимфните синуси, в които лимфата навлиза от стените на тънките черва, са по-широки от тези синуси, които „принадлежат“ на дебелото черво. Този факт също показва по-голямо транспортно натоварване на синусите на "тънкочревната" част на лимфния възел в сравнение с тази на "дебелочревната" част. Това предположение е в съответствие с данните на Н. Рийд (1989), който установи, че тънките черва имат по-голяма абсорбционна способност, а в стените му, голямо количествовещества, отколкото в дебелото черво.
Същото наличие на морфофункционални части се наблюдава в соматичните подколенни лимфни възли, към които тече лимфа от повърхностните и дълбоки тъкани на крайника. Както показва А. И. Краюшкин (1994), лимфата от кожата и подкожна тъкан, а в другата - през дълбоки съдове, от капсулите на ставите, мускулите, сухожилията, фасциите на тазовия крайник на заека. Боята, инжектирана в мускулите и други дълбоко разположени структури на крайника, се изпраща през дълбоките лимфни съдове до подколенния лимфен възел и се намира в синусите на предната част на този възел. Лимфните синуси в задната част на лимфния възел са пълни с боя, която идва от повърхностните тъкани (кожа и подкожна тъкан). Авторът отбелязва, че междинните синуси на медулата са по-широки в тази част на лимфния възел, към която тече лимфа от повърхностните тъкани на крайника. Може да се предположи, че лимфоидният паренхим, съседен на лимфните синуси, който получава лимфа от стените на тънките черва (мезентериални възли) или от кожата и подкожната тъкан (поплитеални възли), ще изпита по-голямо функционално натоварване от паренхима на другата част от тези възли.
Данните, получени от А. И. Краюшкин, потвърждават това предположение. Изследвайки лимфоидната тъкан на лимфните възли, в които се вливат лимфните съдове, носещи лимфа от функционално различни органи, А. И. Краюшкин открива интересни факти. В тази част на лимфния възел, където лимфата тече от стените на тънките черва, лимфоидният паренхим има по-висока количествени показателиотколкото в другата част на възела, свързана с дебелото черво. В "тънкочревната" част на мезентериалния лимфен възел лимфоидните възли (B-зона) са по-големи, диаметърът им варира от 275 до 750 микрона (средно 467 микрона). Площта, заета от лимфоидни възли, е 20,3% от изрязаната площ на мезентериалния лимфен възел. Голяма площ на участъците на възела също е заета от месести нишки. В тези части на мезентериалния лимфен възел има и по-високо съдържание на бластни форми на клетки, големи лимфоцити и плазмоцити, което показва повишаване на лимфоцито- и имунопоетичните функции тук. В "дебелочревната" част на лимфния възел диаметърът на лимфоидните възли е много по-малък -349 μm (200-625 μm), а площта на лимфоидните възли върху участъка на възела е само 13,1%. В тази част на мезентериалния възел, където навлиза лимфата от тънките черва, паракортикалната зона (Т-зоната) заема 3 пъти по-малка площ (8,1%), отколкото в частта, към която тече лимфата от дебелото черво (25,9%) ) .
В соматичния подколенен лимфен възел също има разлики в организацията на лимфоидния паренхим в частите, към които тече лимфата различни регионикрайници. В тази част на възела, към която се приближават лимфните съдове от кожата и подкожната тъкан, има по-високи морфометрични параметри на лимфоидни възли и пулпидни връзки (В-зони). Диаметърът на лимфоидните възли в тази част е 375 μm (193-646 μm), а площта им е 15,2% от цялото сечение на възела. В друга част на подколенния лимфен възел, към който се оттича лимфа от дълбоките тъкани на крайника (мускули, сухожилия, ставни капсули), диаметърът на лимфоидните възли е по-малък от 302 микрона (123-492 микрона), а тяхната площ на среза заема половината от площта -7, 9%. Паракортикалната Т-зона се оказа по-широка в частта, където навлиза лимфата от дълбоките тъкани на крайника.
А. И. Краюшкин обяснява тези разлики в две съседни морфофункционални зони на подколенния лимфен възел с по-силен и постоянен ефект върху кожата на „антигени външна среда". Неслучайно в кожата се откриват периваскуларни лимфоидни възли, които са имунни структури на кожата. Преобладаващото развитие на лимфоидни възли и пулпидни връзки (В-зони) в тази част на централния мезентериален лимфен възел, където лимфата навлиза от тънките черва, и в подколенния възел, към който лимфата се изпраща от кожата и подкожната тъкан, показва формирането на имунни отговори в тези части.хуморален тип. Тези реакции произвеждат антитела. Много изследователи са показали, че лимфоидните възли съдържат имунни комплекси. Според V. I. Novikov и A. A. Vlasov (1989), S. T. Lee, F. Paraskevas и J. Maeba (1985), S. T. Lee и F. Paraskevas (1988), антиген-специфичните комплекси включват антигенни детерминанти, молекули от клас II на хистосъвместимост и имуноглобулин. детерминанти. За образуването на такъв комплекс са необходими макрофаги и Т-лимфоцити.
В. И. Новиков и др. (1990) отбелязват, че клетките на периферните лимфни възли след контакт с антиген in vitro отделят антиген-специфични фактори. Тези фактори потискат миграцията на макрофагите и увеличават тяхната функционална дейност, и също допринасят за крайната диференциация на прогениторни клетки от серията гранулоцити-макрофаги по моноцитния път. Според изследванията на S. K. Drinker, N. Field и N. Ward (1934), именно в лимфните възли може да се открие до 99% от антигенния материал. Следователно в тези възли селективно се натрупват антиген-активирани клетки, които, както показват В. И. Новиков и А. А. Власов (1989), В. И. Новиков, А. А. Власов и И. Г. Сидорович (1991), участват в имунологични реакции срещу този антиген. В пика на имунния отговор клетките на лимфните възли секретират антиген - специфичен хуморален фактор, който засилва генезата на антителата в продуктивния период чрез потискане на функцията на Т-супресорите. Авторите заключават, че клетките на лимфните възли „участват не само в натрупването и преработката на уловения антигенен материал до форма, която с потока на лимфата и кръвта достига до централните органи на имунитета (по-специално, костен мозък), причинявайки наблюдавани промени в производството на имуномодулатори в него, но също така и в регулацията на следващите етапи от развитието на имунния отговор, включително продуктивния период на тази реакция. N. van Rooijen (1987) предполага, че има един начин за миграция на клетки, които се диференцират в лимфните възли в клетки, образуващи антитела. При тимус-независим имунен отговор антиген-реактивните В-клетки получават необходимата информация от антиген-представящите макрофаги в интернодуларната зона на периферната част на кората. M. Dohrzanski и T. Yang (1991) показват, че Т-лимфоцитите на различни лимфни възли се различават по брой и функционални характеристики.
GV Kovalevsky (1992) идентифицира 3 етапа на промени във функционалната морфология на имунния отговор на лимфните възли към тимус-зависими антигени. 1. Преразпределителен стадий - 1 ден от имунния отговор. Антиген-стимулираните регионални възли произвеждат хуморален агент, който насърчава освобождаването на Т-лимфоцити от нерегионални лимфни възли. В същото време в последния се появяват огнища на опустошение на паракортикалната зона на фона на най-острата лимфоцитоза на синусите. В медулата настъпва аутофагия на плазмени клетки и се образуват гигантски сферични макрофаги. 2. Пролиферативен стадий - 3-ти ден от имунния отговор, времето на максимално елиминиране на антигените. Някои области на Т-зоната губят обичайния си дребноклетъчен състав и се превръщат в полета от „големи пиронинофилни клетки“. Предполага се, че всички субпопулации на специфични Т-лимфоцити (помощници, усилватели, супресори, убийци и др.) произлизат от тях. Следователно на този етап се развива авторегулация на имунния отговор в паракортикалната зона. 3. Крайният етап, свързан с морфогенезата, се проявява до 5-6-ия ден от имунния отговор. Има масивна мобилизация на лимфоцити в синусите на регионалните лимфни възли. Рециркулацията включва несенсибилизирани Т-лимфоцити, които допълнително получават антигенна информация чрез контактно взаимодействие (периполеза) със синусови макрофаги. Както отбелязва Г. В. Ковалевски, сенсибилизираните Т-лимфоцити се екстрахират от ендотела на посткапилярните венули на нерегионалните лимфни възли и заселват там местата, възникнали на първия етап. Колкото по-дълго, колкото по-висока е дозата на антигена, се наблюдават "вълни" на репродуктивните центрове.
Според G. G. Csanaky, V. Kalasz и T. Rahr (1991), след антигенна стимулация, както броят на вените с висок ендотел, така и масата на лимфните възли бързо се увеличават.
G. Sainte-Marie и F. S. Peng (1985), T. S. Smirnova (1992) смятат, че клетките, разположени в повърхностната част на кората му (директно под маргиналния синус), допринасят за прехвърлянето на антигени от вливащата се лимфа в паренхима на лимфен възел, наречен маргинална зона. Според G. Sainte-Marie и F. S. Peng (1985) стромата на субкапсуларната зона е представена от фибробластни ретикуларни клетки, образуващи подобна на бримка мрежа. J. E. Veldman, F. J. Heuning и I. Molendar (1978), M. Dobashi, K. Terashima и J. Imai (1982) показват, че в субкапсуларната зона лимфоцитите се трансформират в имунобласти, плазмобласти и плазмени клетки, независимо от Т-клетъчните регулиране и формиране на репродуктивни центрове.
Според Т. С. Смирнова (1992) този процес може да продължи независимо от състоянието на репродуктивните центрове, където развитието и клетъчни трансформациивъзникват много по-бързо, отколкото в други структури на лимфните възли. Т. С. Смирнова също изследва ритмично-времевите промени в популацията на лимфоидните клетки в маргиналната зона и сравнява получените данни с моделите на клетъчния състав в Т- и В-зависимите зони на същия възел. Анализът на резултатите показа липсата на "общи компоненти", което послужи като основа за твърдението за "относителната функционална автономност" на маргиналната зона. Авторът смята, че тази зона е мястото на навлизане в паренхима на лимфния възел на малки лимфоцити и мастоцити от лимфата, навлизаща в възела.
В много изследвания се посочва определен ритъм в структурата на лимфоидните образувания на лимфните възли. В научната литература има доказателства, че масата на соматичните и висцералните лимфни възли се променя през деня. Според Ю. П. Хусар (1969, 1975), максималната маса на лимфните възли на белите плъхове пада на 18 часа, а минималната - на 3 часа. максимална плътностместоположението на лимфоцитите в аксиларните, ингвиналните и мезентериалните лимфни възли също пада на 18 ч. Ю. И. Бородин и др. (1992) разкриват 24-часови биоритми на клетъчния състав в кортикалното плато на ингвиналните лимфни възли на CBA мишки. В кортикалното плато на мезентериалните лимфни възли се наблюдава 12-часов ритъм на промени в броя на лимфоидните клетки: увеличение през нощта и деня, намаляване сутрин и вечер. Авторите обясняват тези факти с двигателната активност по време на хранене през нощта и през деня. Намаляването на броя на клетките в кортикалното плато се случва по време на съня на животните. В това плато на бифуркационни лимфни възли максималното съдържание на клетки се наблюдава през нощта, а минималното - през деня.
Ю. И. Бородин и др. считат, че увеличаването на броя на клетките в нощната фаза е свързано с миграцията на клетките по това време на деня от тимуса, което е в съответствие със заключенията на Ю. П. Хусар (1975). Това не изключва възможността за преразпределение на клетките в самите лимфни възли, както и освобождаването им в кръвния поток при условия на двигателна активност на животните. Става въпрос и за рециркулацията на съществуващите в тялото лимфоцити. Невъзможно е да не се отбележи, както смятат авторите, нестабилността на циркадните ритми в клетъчния състав на кортикалното плато, което зависи от редица причини: позицията на лимфните възли в тялото, нивата на пролиферация, клетките рециркулация и др.
Различен модел на клетъчни ритми е открит в тимус-зависимата паракортикална зона на CBA мишки. В ингвиналните лимфни възли максималният брой клетки от лимфоидната серия пада през деня и вечерта. В паракортикалната зона на мезентериалните лимфни възли се наблюдава намаляване на броя на клетките по време на периода на двигателна активност на животните през нощта. През деня се наблюдава увеличение на броя на клетките и поради двигателната активност. Авторите разглеждат дневната динамика в клетъчния състав във връзка с различни посоки на клетъчна миграция, включващи тимуса, далака, други групи лимфни възли и кръвния поток. Синхронизмът на дневните биоритми на клетъчния състав в кортикалното плато и паракортикалната зона подчертава функционалната връзка на различни лимфоидни структури в тези органи. Според N. Metcalf, S. Youngberger и W. Metcalf (1971) при плъхове скоростта на рециркулация на В-лимфоцитите е по-ниска от тази на Т-лимфоцитите. Според данните на А. Я. Фриденщайн и Е. А. Лурия (1980), в кортикалното вещество, където има много венули и кръвоносни капиляриВ-лимфоцитите мигрират от кръвта през стените на тези микросъдове в паренхима на лимфните възли, в техните лимфоидни възли и месести връзки. В литературата има данни, че до 0,2% от всички В-лимфоцити се намират в тези структури на лимфните възли.
Миграцията на лимфоцитите от лимфоидните възли също се подчинява на законите на дневния ритъм. В лимфоидните възли на бифуркационните лимфни възли се установява 12-часов ритъм на клетъчна активност с увеличаване на броя на клетките в тези структури в сутрешните и вечерните часове, между които има леко намаление на клетките, съдържащи се в тях. "Натрупване на клетки" в лимфоидни възли през деня Ю. И. Бородин и др. (1992) обясняват наличието на митотична активност в тези структури, както и рециркулацията на лимфоцитите. Авторите смятат, че във вечерните часове лимфоцитите мигрират от лимфоидните възли на всички лимфни възли, където и да се намират. Този процес е свързан с началото на вечерната двигателна активност на животните. Двигателната активност е отбелязана както в предзорни часове, така и през деня (хранене на животни). Авторите стигат до извода, че образуването на клетки в лимфоидните възли на лимфните възли се наблюдава по време на съня на животните, а намаляване на техния брой - в периода на физическа активност. При животни, водещи нощен начин на живот, процесът на миграция на лимфоцити от лимфоидни възли естествено се случва през нощта.
В кашестите нишки на соматичните (ингвинални) и висцералните (бифуркационни и мезентериални) лимфни възли се наблюдава 12-часов ритъм на клетъчна активност. Сутрин и вечер броят на клетките от лимфоидната серия в пулпните връзки намалява, а през деня и през нощта се увеличава, което не е в съответствие с биоритмите на клетките в лимфоидните възли. Следователно Ю. И. Бородин и др. (1992) заключават, че пулпните връзки не са напълно B-зависими структури на лимфните възли. Динамиката на клетките в пулпизните струни е по-подобна на същите процеси в кортикалното плато и паракортикалната зона и авторите предполагат акумулативна роля на пулпизните струни по отношение на клетките от лимфоидната серия. Биотехнологично изследване на функционалните характеристики на пулпните връзки разкрива връзката им с В- и Т-зоните на лимфните възли. Притокът на лимфоцити в пулпните връзки отвън се отбелязва през първата половина на деня, а имиграцията - през нощта, когато физическа дейностживотни е много висока.
Що се отнася до биоритмите в лимфоидните структури на други органи на имунната система, Yu.P.Hussar, Yu.Teikes и X.Lyaene (1971), L.Scheving et al. (1972) показват промени в масата на далака при плъхове през деня. Увеличаване на масата се наблюдава по време на почивка, максималната маса се отбелязва преди зазоряване, а минималната - през деня и вечерта.
Приблизително същата картина на увеличаване на броя на клетките на лимфоидната серия сутрин и намаляване на вечерните и дневните часове се наблюдава от Ю. П. Хусар (1975) в бялата пулпа на далака при мъжки CBA мишки . Значителното увеличение на броя на лимфоцитите и увеличаването на плътността на тяхното местоположение в бялата пулпа на далака може да се дължи на миграцията на тези клетки от костния мозък, където по това време броят на тези клетки намалява. През нощта също се отбелязва повишаване на митотичната активност в далака.
Възможно е увеличаването на масата на лимфоидната тъкан в далака в ранните сутрешни часове и през втората половина на дневните часове да се дължи на активното преразпределение на лимфоидните клетки в условията на промени във функционалните натоварвания на различни органи по време на ден. Може да се предположи, че през деня лимфоцитите са по-заети с контрола на хранителните антигени в стените на храносмилателните органи и в други органи, които активно работят през деня. Както пишат Ю. И. Бородин и др. (1992), във вечерните и нощните часове "експертната система" регистрира освобождаването на клетки от далака в кръвта и В-зависимите зони на лимфните възли. Възможно е "контролът" на кръвта, който се извършва през всичките 24 часа в денонощието, да достигне максимум в предсутрешните часове. Както е показано от G. Fernandes et al. (1976), през деня в далака има минимален брой Т-лимфоцити с едновременно максимално съдържание в периферната кръв.
Експериментите, проведени от Т. С. Смирнова с нанасяне на наранявания на животни, също показват значителни промени в клетъчния състав на лимфните възли (в тяхната маргинална зона). След нараняването (на 1-2-ия ден) в тази област броят на малките лимфоцити се е увеличил със 75%. Това увеличение в l "/ 2 пъти е по-голямо, отколкото в други области на лимфните възли. Т. С. Смирнова заключава, че лимфоцитите идват в маргиналната зона от мястото на нараняване. Авторът не обяснява откъде идват лимфоцитите в областта на нараняване и защо именно оттам те навлизат в лимфния възел.Както посочва Т. С. Смирнова, до 7-8-ия ден след нараняването "гъстотата на популацията" на лимфоцитите в маргиналната зона се нормализира.В същото време тя обръща внимание на различното поведение на други клетки от лимфоидната серия, броят на които в маргиналната зона не се увеличава, а напротив, намалява. По този начин до 3-ия ден след нараняването броят на средните лимфоцити намалява с повече от 3 пъти, митотично делящи се клетки - 6 пъти, бластни форми на клетки - 21/2 пъти, незрели плазмени клетки - 3 пъти, зрели плазмени клетки - 10 пъти Т. С. Смирнова също обръща внимание на наличието в посттравматичните период в маргиналната зона на регионалния лимфен възел на мастоцитите, около който рязко нараства към 3-ия ден. Броят на мастоцитите също се увеличава в зоната на нараняване. Този синхрон на появата на мастоцитите на мястото на нараняване и маргиналната зона показва, според автора, възможно увеличаване на интензивността на локалния кръвен поток, стимулиране на фагоцитната активност на макрофагите и наличието на хемотаксис в лимфоцитите. . В същото време Т. С. Смирнова се позовава на данните на Н. А. Юрина и А. И. Радостина (1977) за ролята на мастоцитите в организма и J. Raud et al. (1989) относно периартериалното местоположение на мастоцитите, за да помогне за насочване на миграцията на левкоцитите. При травма в маргиналната зона се променя и ритъмът на клетъчната популация. Вярно е, че амплитудата на колебанията и промените, идентифицирани от Т. С. Смирнова след нараняването, намалява.
Въз основа на проведените изследвания Т. С. Смирнова смята, че във функционален план маргиналната зона заема специално място в лимфния възел. Авторът смята, че маргиналната зона е мястото на навлизане на "лимфоцити и мастоцити в паренхима на лимфните възли от лимфния канал, както при нормални условия, така и в отговор на хирургична травма". По този начин, на настоящия етап от морфологичните изследвания, хронобиологичните наблюдения са много обещаващи. Те правят възможно по-доброто разбиране на моделите на морфогенетичните процеси на клетъчно, тъканно и органно ниво. По отношение на лимфните възли също се оказа (в експеримента), че дневните биоритми на кръвоснабдяването на лимфоидната тъкан са в пряко съответствие с дневните ритми на кръвоснабдяването на тъканните области. Локалните промени в лимфните възли се влияят от хормонални и нервни фактори, динамиката на ежедневната популация на мастоцитите. Възможно е мастоцитите, присъстващи в периваскуларните и лимфоидните тъкани, в синусите, капсулата и трабекулите на лимфните възли да повлияят както на микросредата, така и на формирането на имунния отговор. Мастните клетки могат не само да образуват групи от 3-5 елемента, но дори и "гъбовидни" структури, които започват в областта на портата на възела и продължават по трабекулите и съдовете до кортикалното междувъзлово плато. Подробностите за усвояването на освободените мастоцитни гранули от ретикуларните клетки все още не са ясни. Авторът смята, че маргиналната зона е мястото на навлизане в паренхима на лимфния възел на малки лимфоцити и мастоцити от лимфата, навлизаща в възела.
Полагането на първите лимфни възли е установено при 5-6-седмични човешки ембриони. Според V. A. Trufakin (1967), първите мезентериални лимфни възли започват да се образуват на 8-та седмица от вътрематочното развитие. V. A. Florensov (1983) установи, че най-ранното полагане на лимфните възли се наблюдава вече при 7-седмични ембриони. Анатомично оформените лимфни възли стават при 9-седмични фетуси. М. А. Долгова (1967), Н. А. Жарикова (1979) и други изследователи посочват тази възраст (8-10 седмици), на която започва образуването на лимфни възли. Трябва да се отбележи, че времето на образуване на лимфни възли, според различни автори, като правило, е различно. Това може да се разбере, като се има предвид следното. Първо, различни групи лимфни възли често са обект на изследване. Второ, не е взето предвид здравето на майката, която е носила тези ембриони и фетуси, условията за получаване на материала и т. н. В това отношение трябва да се съгласите с G. V. Kharlova (1963), че образуването на лимфни възли е компенсаторен отговор на условията на съществуване на организма.
Тъй като лимфните възли са разположени по пътищата на лимфния поток от органи и тъкани, ранното появяване на възли не е изненадващо. Още през първите седмици от развитието на ембриона в тъканната течност се появяват метаболитни продукти, включително груби протеини и дори частици от разрушени клетки, образувани в резултат на обновяването на клетките. Тъй като мъртвите клетки, грубите протеини и възможните мутантни клетки са чужди частици, за тяхното разпознаване и унищожаване са необходими имунни структури. Такива структури са лимфните възли, образувани в тялото на ембриона, които са биологични филтри по пътищата на движение на течността към кръвния поток. Вероятно появата в тялото на ембриона на метаболитни продукти и чужди частици в количество, когато стане опасно за растящия организъм, е стимулът, отправната точка за образуването на лимфни възли. В различни области на тялото се създава такава необходимост от появата на лимфни възли и техните регионални групи в различно време. Следователно появата на раздели на възли в различни областичовешкото тяло възниква в различни периоди на вътрематочно развитие, до раждането и дори след него.
Образуването на лимфен възел започва с изолирането на група мезенхимни клетки в близост до лимфен съд или между съседни лимфни съдове. След това, в процеса на развитие, клетъчният клъстер се инвагинира в лумена на съседния лимфен съд, чиято външна стена дава начало на капсулата на получения лимфен възел. Луменът на лимфния съд допълнително се превръща в субкапсуларен (маргинален) синус. Междинните синуси се развиват на базата на разклонен плексус от лимфни съдове, между които растат нишки от ембрионална съединителна тъкан. В бъдеще броят на мезенхимните клетки в рудимента на възела се увеличава. На 3-4-ия месец от вътрематочното развитие се определят лимфоцити в развиващите се лимфни възли.
A. Yasushi (1992) установи, че при фетусите лимфоидните възли започват да се появяват на 16-та седмица от развитието под субкапсуларния синус. От 18-та седмица постоянно се откриват добре видими лимфни възли. Авторът обяснява липсата на репродуктивни центрове в нодулите с незрялостта на тъканите и нодуларните дендритни клетки, както и с липсата на стимулация по време на ембрионалното развитие. Според J. Westerga и W. Timens (1989) В-клетките със среден размер се откриват в лимфоидните възли и външните участъци на кортикалната субстанция на лимфните възли при фетуси. Започвайки от 19-та седмица, в някои възли може да се види очертаната граница между бъдещата кортикална и медула. По този начин лимфоидните възли в лимфните възли се появяват още в пренаталния период. P. Fusari (1965) наблюдава образуването на възли при 51/2-месечен плод, V. A. Florensov (1964) - на 34-та седмица от вътрематочното развитие. Центровете за възпроизвеждане в лимфоидните възли се появяват малко преди раждането или малко след него.
Диференциацията на лимфните възли продължава не само при новородени, но и при детство. Основните свързани с възрастта процеси на формиране в лимфните възли завършват до 10-12-годишна възраст, но преструктурирането на тези възли се извършва през целия живот в съответствие с променящите се условия на съществуване. Свързаната с възрастта инволюция на лимфните възли може да се наблюдава от юношеството. С напредването на възрастта техният брой намалява и средният размер на останалите възли се увеличава. Това е така, защото много малки лимфни възли са напълно заменени от плътна фиброзна съединителна и мастна тъкан. Близките възли растат заедно и образуват по-големи възли със сегментна или лентовидна форма. От юношеството групи от мастни клетки се появяват в стромата и паренхима на лимфните възли. С напредването на възрастта количеството на съединителната тъкан в възлите се увеличава, капсулата се удебелява и хиларните трабекули нарастват значително; влакната на ретикуларната строма се удебеляват. Плътната фиброзна съединителна и мастна тъкан с напредване на възрастта постепенно измества лимфоидния паренхим в лимфните възли. В същото време количеството на кортикалното вещество в възлите намалява и съдържанието на медулата се увеличава относително. Междинните синуси в медулата се разширяват. Има и възрастови особености на клетъчния състав на лимфните възли; броят на малките лимфоцити и макрофагите се увеличава, докато броят на средните лимфоцити намалява.